Research Article |
|
Corresponding author: Xiao Guo ( xiaoguoyeah@yeah.net ) Academic editor: Graeme Bourdôt
© 2023 Xiao Guo, Xin-Yue Liu, Si-Yu Jiang, Shao-Xia Guo, Jing-Feng Wang, Yi Hu, Shi-Mei Li, Hai-Mei Li, Tong Wang, Ying-Kun Sun, Ming-Yan Li.
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Guo X, Liu X-Y, Jiang S-Y, Guo S-X, Wang J-F, Hu Y, Li S-M, Li H-M, Wang T, Sun Y-K, Li M-Y (2023) Allelopathy and arbuscular mycorrhizal fungi interactions shape plant invasion outcomes. NeoBiota 89: 187-207. https://doi.org/10.3897/neobiota.89.110737
|
Abstract
The novel weapon hypothesis suggests that allelopathy is an important mechanism for exotic plants to successfully invade native plant communities. Allelochemicals from exotic plants affect both native plants and arbuscular mycorrhizal fungi (AMF) in soil. To explore these effects, we conducted pot experiments using a native plant community comprising of Chenopodium album L., Vitex negundo L., Rhus chinensis Mill., and Acer truncatum Bunge. We incorporated AMF strains (Funneliformis mosseae, Glomus versiforme, and Rhizophagus intraradices in a 1:1:1 volume ratio) into a soil mixture comprising equal volumes of autoclaved field soil and grass ash at five concentrations (0 g L–1, CK; 10 g L–1, LRE; 20 g L–1, NRE; 30 g L–1, MRE; and 40 g L–1, HRE) to investigate the allelopathy of Rhus typhina L. root and its interactions with AMF on the native plant community. Our results indicated that low concentrations of allelochemicals promoted the relative growth rate and leaf nitrogen content in the native plant community, which was attributed to the increased environmental stress that improved the degree of leaf photosynthetic capacity and organic synthesis rate in the native communities. Moreover, allelochemicals with toxicity decreased the colonization rates of AMF. Meanwhile, the presence of allelopathic effects induced a decrease in leaf nitrogen, and allelopathy altered the effects of AMF on the native community. Specifically, the high concentration of allelochemicals altered the positive effect of AMF on the total aboveground biomass of the native plant community to a negative effect, likely by decreasing colonization rates or affecting soil physicochemical properties and the composition of the mycorrhizal communities. Consequently, it is necessary to consider the effects of AMF when testing the roles of allelopathy or the novel weapons hypothesis in biological invasions.
Graphical abstract
Keywords
Allelochemicals, mycorrhizal symbionts, photosynthetic acquisition capacity, physiological and growth parameters
Introduction
Exotic invaders are a worldwide problem, which can displace native species, causing significant changes in the diversity of native communities (
Allelopathic effects refer to the positive or negative interaction between plants mediated by chemical compounds (
Arbuscular mycorrhizal fungi (AMF), a representative group of the mycorrhizal communities, can form symbiotic associations with approximately 70% of plant species (
Invasive plants can produce distinct allelochemicals, which can disturb the structure of mycorrhizal communities in the rhizosphere and alter soil nutrient cycling patterns (
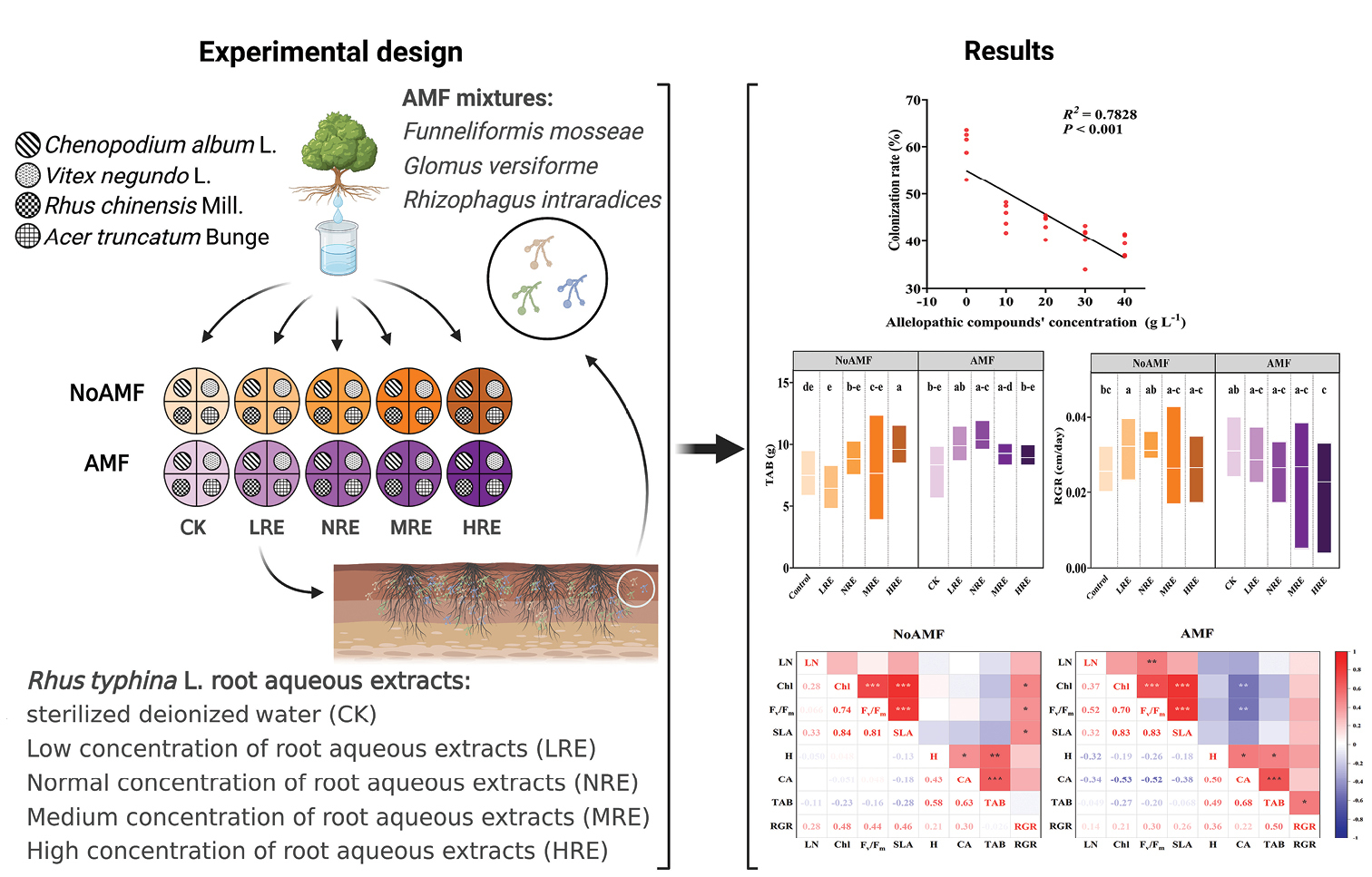
In this study, we selected four common plants of the warm temperate zone of China, namely Chenopodium album L., Vitex negundo L., Rhus chinensis Mill., and Acer truncatum Bunge, to simulate a native plant community. We used root aqueous extracts of Rhus typhina L. to simulate allelopathic effects of invasive plants, and inoculated AMF (Funneliformis mosseae, Glomus versiforme, Rhizophagus intraradices). We conducted a community-level greenhouse experiment to test the following hypotheses: 1) the allelopathic effects of R. typhina could negatively impact the growth of the native plant community, 2) the allelopathic effects of R. typhina could negatively impact the colonization rates of AMF, and 3) the allelopathic effects of R. typhina could alter the impact of AMF on the growth of the native plant community.
Materials and methods
Study species
Rhus typhina L., an exotic tree species indigenous to North America, was introduced to China as a horticultural and greening ornamental plant in 1959 (
In mid-April 2021, we prepared sufficient quantities of seeds of the three selected plant species. The following treatments were performed to improve their germination rate. First, seeds of R. chinensis were treated with a baking soda solution at 70 °C for 10 minutes to remove their waxy skins and subsequently cleaning with sterile deionized water. Second, seeds of A. truncatum were subjected to a 4 °C treatment for 24 hours. Third, seeds of both R. chinensis and A. truncatum, as well as those of V. negundo, were soaked in fresh water for 24 hours, replacing the water every 12 hours. Soaked seeds were then evenly spread into trays with three layers of gauze on the bottom, and two layers of gauze were placed over the seed surface to improve germination. During this process, sterile deionized water was sprayed into the trays daily to maintain moisture levels. After approximately 30% of the seeds had germinated, we selected strong and uniform seedlings of each species and transferred them to cell trays for further culture. Finally, seedlings of C. album with 4–5 leaves were collected from the campus of Qingdao Agricultural University, Shandong, China, until three plant seedlings had developed 4–5 leaves.
Preparation of allelopathic solutions of R. typhina
There are two primary rationales for using the root aqueous extracts from R. typhina to study the allelopathic effects: (1) allelopathic effects of R. typhina are mainly mediated by phenolic substances, most of which, especially polyphenols, can be dissolved in water (
Preparation of AMF
The AMF inoculum comprised a blend of three common AMF strains – Funneliformis mosseae, Glomus versiforme, and Rhizophagus intraradices – in a 1:1:1 volume ratio. As different AMF types have varying colonization rates in diverse plant species, the AMF mixtures can ensure consistent colonization rates across diverse plant species. In the AMF treatments, each pot was inoculated with the AMF mixtures containing approximately 8 × 103 viable propagules. In the NoAMF treatments, an equivalent amount of sterilized AMF inoculum was added to prevent substrate effects.
Experimental design
The experiment was performed at Qingdao Agricultural University, situated in Qingdao, China (36°31'N, 120°39'E). The region has a temperate monsoon climate with an annual temperature of approximately 12.7 °C, mean annual precipitation of around 821.8 mm, and average yearly sunshine duration of about 2,541.1 h. In the summer of 2021, we conducted a 70-day experiment in an artificially-controlled greenhouse maintained at an average temperature of 29.4 °C and a relative humidity of 77%, using a digital display humidity thermometer, DL-WS20 (Hangzhou Gsome Technology Co., China). The plastic pots used in this study were 20.0 cm in height and 20.0 cm in diameter, and were filled with soil materials consisting of autoclaved field soil and grass ash mixed at a 1:1 volume ratio.
In each plastic pot, we planted four different species of seedlings (C. album, V. negundo, R. chinensis, and A. truncatum), with four to five leaves per seedling. One seedling for each plant species was transplanted into one of the four equal sections of the cross lines past the center of the circle within each pot, resulting in a total of four plants per pot. The control treatment consisted of no AMF and root aqueous extracts of R. typhina. Overall, we established 100 experimental pots, including two AMF treatments (with and without AMF) and five concentrations (0 g L– 1, CK; 10 g L– 1, LRE; 20 g L– 1, NRE; 30 g L– 1, MRE; and 40 g L– 1, HRE). Each treatment had ten replicates (n = 10), and all individuals of each species and treatments were randomly distributed. Over the entire experiment, we added 50 ml of the appropriate concentration solution of the root aqueous extracts from R. typhina once every two days in the five concentration treatments, for a total of thirty times. Weeding and pest control measures were implemented as usual and the greenhouse was adequately ventilated. All pots were placed randomly within the greenhouse and rotated normally to minimize any potential effects of environmental differences during the experiment.
Harvest and measurements
Physiological parameters measured in the native plant community included specific leaf area (SLA), total chlorophyll concentration (Chl), Fv/Fm, and leaf nitrogen (LN), which can characterize the degree of leaf photosynthetic capacity (
Prior to the treatment, the aboveground component of five seedlings from each indigenous species with 4–5 true leaves were oven-dried at 105 °C for 0.5 h to inactivate them. The drying process was continued for an additional 48 h at 80 °C, and then the weight was measured. The mean value was regarded as the initial biomass for each species.
After 60 days, when the herbaceous flora had reached the reproductive stage, the maximum growth index of all native plants was determined. The vertical height of the plant from the base to the terminal bud was gauged as the plant height. The CA was calculated using the diamond-shaped area formula (
CA = 0.5 × a × b
where a indicates the length of the plant’s maximum horizontal extension, and b signifies the length perpendicular to a on the plant’s maximum horizontal extension plane.
For each plant, chlorophyll fluorescence parameters were evaluated during sunny and cloudless weather conditions from 8:30 to 11:30 using Pocket PEA (Hansatech Instruments Ltd., UK). The top-to-bottom first fully developed leaf was selected, and after undergoing dark treatment for 0.5 h, the maximum fluorescence value (Fm) and variable fluorescence value (Fv) were evaluated. The maximum photochemical efficiency (Fv/Fm) of PSII was then calculated (
The second or third fully expanded and healthy leaf from the apical meristem of each plant was selected, cleaned with sterilized deionized water, and the leaf area was measured using a portable leaf area meter (Yaxin-1241, Yaxin Inc., Beijing, China). The leaves were inactivated to inactivation at 105 °C for 0.5 h and dried at 85 °C for 24 h in a drying oven until they reached constant weight to obtain their dry weights. The SLA was calculated using the formula (
SLA = leaf area ÷ dry weight
The total chlorophyll content was determined using the ethanol extraction method. Four fully mature leaves near the top of each plant (one leaf per plant) were collected for each treatment and chopped to approximately 0.2 g fresh weight. The leaves were then submerged in 10 ml of 95% alcohol until completely whitened. The absorbance of chlorophyll in the solution at 665 nm and 649 nm wavelengths was measured using a UH5300 UV/VIS spectrophotometer (Hitachi, Inc., Tokyo, Japan). The concentrations of chlorophyll a, b, and total chlorophyll were calculated using the following formulas (
Chlorophyll a concentration = 13.95 × A665 – 6.88 × A649
Chlorophyll b concentration = 24.96 × A649 – 7.32 × A665
Chlorophyll content
= Chlorophyll concentration
× volume of extraction solution / fresh weight of sample
Chl = Chlorophyll a content + Chlorophyll b content
The aboveground parts of the plants were harvested, washed, classified according to species, inactivated at 105 °C for 0.5 h in the oven, and dried in the oven at 80 °C for 48 h. The total aboveground biomass of all species in the same pot was measured and the RGR of native plants was calculated using the formula (
RGR = (ln X1 – ln X2) ÷ ∆T
where X2 and X1 denote the final and initial biomass, respectively, and ∆T is the duration of the experiment.
For LN measurements, dried leaves of each species were ground and weighed to 0.5 g using the Kai-style digestion method (K9860, Hanon, Shandong, China).
Roots were collected from 70-day plants. Fine roots (15 g) were excised from each plant, washed, and stained with acid fuchsin to determine the percentage of mycorrhizal colonization by the method of Biermann and Linderman using a BX50 Olympus microscope (Olympus Optical Co., Ltd., Tokyo, Japan) (
Data analyses
The Community-Weighted Trait (CWT) method was utilized to evaluate the relevant physiological parameters (leaf nitrogen, total chlorophyll concentration, Fv/Fm, and specific leaf area) and growth parameters (height, crown area, and relative growth rate) of the native plant community in this study (
where S is the total number of species and Pi represents the relative abundance of species i in the native community. Pi = ni ÷ N, ni and N represent the plant number of species i and the number of plants of all species in the native plant community, separately. Xi denotes the average value of morphological and physiological characteristics of species i.
To assess differences in growth and physiological traits among different treatments, we conducted a two-way analysis of variance (ANOVA). Prior to the ANOVA, we performed variance homogeneity tests for each group of data and transformed unevenly distributed data. We used Duncan’s test with a significance level of α ≤ 0.05. For each concentration of allelochemicals, we performed linear regressions to determine the relationship between AMF colonization rates and the concentration of root aqueous extracts from R. typhina. To evaluate the correlation level of the main traits for the native plant community, we generated Pearson product-moment correlation coefficients among the physiological and growth parameters. We used the IBM SPSS Statistics 25.0 software package (IBM Corporation, Armonk, NY, USA) to conduct all analyses. We created all figures using Origin 2021 (OriginLab Co., Northampton, MA, USA).
To gain an understanding of how growth and physiological parameters of the native plant community were influenced by allelopathy and AMF, and how these parameters were moderated, structural equation modeling (SEM) was utilized. A conceptual model, based on theoretical interactions among variables, was established to serve as our formal hypothesis. To test our hypotheses statistically, the variance-covariance matrix implied by the conceptual model was compared with the observed variance-covariance matrix, and maximum likelihood estimation was employed to estimate model coefficients. Model fit was considered acceptable if the χ2 test was non-significant, the root mean square error of approximation (RMSEA) was low (RMSEA < 0.01), and the comparative fit index (CFI) was high (CFI > 0.90). SEM was conducted using the lavaan and semPlot package in R 4.2.3 (R Core Team 2023).
Results
Plant physiology
Physiological parameters such as LN and Chl were significantly affected by allelopathy (p < 0.001, Table
Results of two-way ANOVA for the effects of allelopathy, AMF and their interaction on growth and physiological parameters of native plant community. CA, crown area; RGR, relative growth rate; TAB, total aboveground biomass; LN, leaf nitrogen; Chl, total chlorophyll concentration; SLA, specific leaf area. Data are presented with F value (n = 10). Significant effects are indicated by bold font (p ≤ 0.05).
| Growth parameters | Physiological parameters | |||||||
|---|---|---|---|---|---|---|---|---|
| CA | Height | RGR | TAB | LN | Chl | Fv/Fm | SLA | |
| Allelopathy | 1.274 | 2.109 | 2.567 | 2.183 | 34.661 | 2.671 | 0.257 | 1.208 |
| AMF | 1.011 | 2.259 | 1.041 | 3.589 | 241.722 | 1.921 | 3.662 | 0.004 |
| Allelopathy × AMF | 0.283 | 2.288 | 2.254 | 4.716 | 24.352 | 1.667 | 1.172 | 0.702 |
The AMF inoculation significantly affected only LN, which was depressed within the LRE, NRE, MRE, and HRE treatments (p < 0.001, Table

Responses of AMF colonization rate to the different concentrations of allelochemicals (n = 5). Five different concentrations of Rhus typhina L. root aqueous extracts were used, which were 0, 10, 20, 30, and 40 g L- 1, respectively. The proportion of variance is explained by R2. The regression line was significant at the p < 0.05 level.

Responses of physiological parameters of native plant community to the allelopathy and its interactions with AMF (n = 10). The boxes represent the range of maximum and minimum values, with a bold horizontal line indicating the average value. Colors indicate different AMF treatments (orange, AMF; purple, NoAMF). The physiological parameters include leaf nitrogen (LN) (A) total chlorophyll concentration (Chl) (B) Fv/Fm (C) and specific leaf area (SLA) (D). There were five different concentrations of the root aqueous extracts of Rhus typhina including 0 g L− 1 (CK), 10 g L− 1 (LRE), 20 g L− 1 (NRE), 30 g L− 1 (MRE), and 40 g L– 1 (HRE). The different letters indicate a significant difference (p ≤ 0.05) with Duncan’s test.
Most physiological trait values were not influenced by the interaction between allelopathic effects and AMF treatments, such as Chl, Fv/Fm, and SLA (Table
Plant growth
The two-way ANOVA indicated that only RGR was altered by allelopathic effects (p = 0.043, Table

Responses of the growth parameters of native plant community to the allelopathy and its interactions with AMF (n = 10). The boxes represent the range of maximum and minimum values, with a bold horizontal line indicating the average value. Colors indicate different AMF treatments (orange, AMF; purple, without AMF). The growth parameters include height (A) crown area (CA) (B) total aboveground biomass (TAB) (C) and relative growth rate (RGR) (D). There were five different concentrations of the root aqueous extracts of Rhus typhina including 0 g L− 1 (CK), 10 g L− 1 (LRE), 20 g L− 1 (NRE), 30 g L− 1 (MRE), and 40 g L– 1 (HRE). The different letters indicate a significant difference (p ≤ 0.05) with Duncan’s test.
Inoculation with AMF increased TAB by 57% in the LRE treatment and decreased it by 29% in the HRE treatment compared to the NoAMF treatment (p = 0.006, Table
The interaction between allelopathic effects and AMF significantly affected on TAB and the LRE and NRE treatments increased TAB by 48% and 45% respectively, compared to the control (p = 0.005, Table
Trait correlation
We examined 28 pairs of growth and physiological traits in the native community using a Pearson correlation test. Nine pairs for the NoAMF treatments and 10 pairs for the AMF treatments were significantly correlated at p ≤ 0.05 (Fig.

Matrix of Pearson product-moment correlation coefficients for the growth and physiological traits of the native community across different AMF treatments (n = 10). Traits include leaf nitrogen (LN), total chlorophyll concentration (Chl), Fv/Fm, specific leaf area (SLA), plant height (H), crown area (CA), total aboveground biomass (TAB), and relative growth rate (RGR). Significant correlations are denoted by bold font and asterisks: *** P ≤ 0.001, ** P ≤ 0.01, and * P ≤ 0.05.
The SEMs were well-ftted to the data (χ2 = 4.507, df = 11, P = 0.953, CFI = 1.000, RMSEA = 0.000). Allelopathy had direct negative associations with relative growth rate, total chlorophyll concentration, specific leaf area, leaf nitrogen and Fv/Fm but positive associations with height, crown area and total aboveground biomass (Fig.

The structural equation models relating the growth and physiological parameters of the native plant community. Green lines indicate positive relationships while red lines indicate negative relationships. Line thickness depicts the magnitude of path coefficients. Significant effects are indicated by asterisks: *** P ≤ 0.001, ** P ≤ 0.01, and * P ≤ 0.05.
Mycorrhizal colonization
The colonization rates of mycorrhizal in the native plant community were negatively correlated with the concentrations of root aqueous extracts from R. typhina (p < 0.001; R2 = 0.7828, Fig.
Discussion
The allelopathy and its interactions with AMF affected the physiological and growth parameters of the native plant community. In general, the low concentration of allelochemicals promoted RGR and LN in the native plant community, while the present of allelochemicals altered AMF effects on the native community.
Responses of native plant community to allelopathy
Allelopathy, as a plants’ defense mechanism, could alter the physical and chemical properties of the soil conditions (
Allelochemicals have long been considered as a mechanism by which invasive species eliminate natives (
Additionally, we found that the total aboveground biomass of the native plant community was significantly higher in the HRE treatment than in the other extract treatments. We analyzed the allelopathic effects on the aboveground biomass of four species (C. album, A. truncatum, R. chinensis, and V. negundo) that constituted the native plant community separately. Only the aboveground biomass of C. album increased significantly with the HRE treatment, while the aboveground biomass of the remaining three plants remained unchanged statistically. A previous study suggests that under the presence of allelopathic compounds, distinct plant species exhibited varying germination and growth rates (
Responses of the native plant community to AMF and allelopathy
AMF can facilitate host plants growth and provide them with competitive advantages (
While the significance of AMF alone in soil has long been recognized in relation to plant invasions, few studies have examined the influence of AMF on allelopathic effects in the context of communities (
The high concentration of allelochemicals transformed the promotive effect of AMF on TAB into the inhibitory effect. Our results suggest that the high concentration of allelochemicals had a neutral impact on the native plant community. Therefore, this phenomenon can be attributed to two plausible reasons: (1) the allelopathic effects from invasive plants can influence patterns of soil nutrient cycling and the soil nutrient environment, altering the effect of AMF on the host (
Conclusion
As biological invasions continue to worsen, it is increasingly necessary to explore the mechanisms behind successful species invasions (
Acknowledgements
The research was supported by National Natural Science Foundation of China (No. 32271588), Science & Technology Specific Projects in Agricultural High-tech Industrial Demonstration Area of the Yellow River Delta (No. 2022SZX16) and Talent Introduction Program for Youth Innovation Team of Shandong Higher Learning (No. 018-1622001).
References
- Adomako MO, Ning L, Tang M, Du DL, van Kleunen M, Yu FH (2019) Diversity- and density-mediated allelopathic effects of resident plant communities on invasion by an exotic plant. Plant and Soil 440(1–2): 581–592. https://doi.org/10.1007/s11104-019-04123-9
- Agren GI, Franklin O (2003) Root: Shoot ratios, optimization and nitrogen productivity. Annals of Botany 92(6): 795–800. https://doi.org/10.1093/aob/mcg203
- Antunes PM, Miller J, Carvalho LM, Klironomos JN, Newman JA (2008) Even after death the endophytic fungus of Schedonorus phoenixreduces the arbuscular mycorrhizas of other plants. Functional Ecology 22(5): 912–918. https://doi.org/10.1111/j.1365-2435.2008.01432.x
- Bais HP, Vepachedu R, Gilroy S, Callaway RM, Vivanco JM (2003) Allelopathy and exotic plant invasion: From molecules and genes to species interactions. Science 301(5638): 1377–1380. https://doi.org/10.1126/science.1083245
- Bennett JA, Klironomos J (2019) Mechanisms of plant-soil feedback: Interactions among biotic and abiotic drivers. The New Phytologist 222(1): 91–96. https://doi.org/10.1111/nph.15603
- Biermann B, Linderman RG (1981) Quantifying vesicular-arbuscular Mycorrhizae: A proposed method towards standardization. The New Phytologist 87(1): 63–67. https://doi.org/10.1111/j.1469-8137.1981.tb01690.x
- Callaway RM, Ridenour WM (2004) Novel weapons: Invasive success and the evolution of increased competitive ability. Frontiers in Ecology and the Environment 2(8): 436–443. https://doi.org/10.1890/1540-9295(2004)002[0436:NWISAT]2.0.CO;2
- Callaway RM, Cipollini D, Barto K, Thelen GC, Hallett SG, Prati D, Stinson K, Klironomos J (2008) Novel weapons: Invasive plant suppresses fungal mutualists in America but not in its native Europe. Ecology 89(4): 1043–1055. https://doi.org/10.1890/07-0370.1
- Callaway RM, Bedmar EJ, Reinhart KO, Silvan CG, Klironomos J (2011) Effects of soil biota from different ranges on Robinia invasion: Acquiring mutualists and escaping pathogens. Ecology 92(5): 1027–1035. https://doi.org/10.1890/10-0089.1
- Chen E, Liao H, Chen B, Peng S (2020) Arbuscular mycorrhizal fungi are a double-edged sword in plant invasion controlled by phosphorus concentration. The New Phytologist 226(2): 295–300. https://doi.org/10.1111/nph.16359
- Cheng F, Cheng Z (2015) Research Progress on the use of Plant Allelopathy in Agriculture and the Physiological and Ecological Mechanisms of Allelopathy. Frontiers in Plant Science 6: 1020. https://doi.org/10.3389/fpls.2015.01020
- de Vries FT, Griffiths RI, Knight CG, Nicolitch O, Williams A (2020) Harnessing rhizosphere microbiomes fordrought-resilient crop production. Science 368(6488): 270–274. https://doi.org/10.1126/science.aaz5192
- Delavaux CS, Smith-Ramesh LM, Kuebbing SE (2017) Beyond nutrients: A meta-analysis of the diverse effects of arbuscular mycorrhizal fungi on plants and soils. Ecology 98(8): 2111–2119. https://doi.org/10.1002/ecy.1892
- Djurdjević L, Gajić G, Kostić O, Jarić S, Pavlović M, Mitrović M, Pavlović P (2012) Seasonal dynamics of allelopathically significant phenolic compounds in globally successful invader Conyza canadensis L. plants and associated sandy soil. Flora – Morphology, Distribution. Flora (Jena) 207(11): 812–820. https://doi.org/10.1016/j.flora.2012.09.006
- Dong LJ, Ma LN, He WM (2021) Arbuscular mycorrhizal fungi help explain invasion success of Solidago canadensis. Applied Soil Ecology 157: 103763. https://doi.org/10.1016/j.apsoil.2020.103763
- Gross EM (2003) Allelopathy of aquatic autotrophs. Critical Reviews in Plant Sciences 22(3–4): 313–339. https://doi.org/10.1080/713610859
- Gruntman M, Zieger S, Tielbörger K (2016) Invasive success and the evolution of enhanced weaponry. Oikos 125(1): 59–65. https://doi.org/10.1111/oik.02109
- Guo X, Hu Y, Ma J-Y, Wang H, Wang K-L, Wang T, Jiang S-Y, Jiao JB, Sun Y-K, Jiang X-L, Li M-Y (2023a) Nitrogen Deposition Effects on Invasive and Native Plant Competition: Implications for Future Invasions. Ecotoxicology and Environmental Safety 259: 115029. https://doi.org/10.1016/j.ecoenv.2023.115029
- Guo X, Ma J-Y, Liu L-L, Li M-Y, Wang H, Sun Y-K, Wang T, Wang K-L, Meyerson LA (2023b) Effects of salt stress on interspecific competition between an invasive alien plant Oenothera biennis and three native species. Frontiers in Plant Science 14: 1144511. https://doi.org/10.3389/fpls.2023.1144511
- Hassan MO, Mohamed HY (2020) Allelopathic interference of the exotic naturalized Paspalum dilatatum Poir. threatens diversity of native plants in urban gardens. Flora (Jena) 266: 151593. https://doi.org/10.1016/j.flora.2020.151593
- Hawkes CV, Belnap J, D’Antonio C, Firestone MK (2006) Arbuscular Mycorrhizal Assemblages in Native Plant Roots Change in the Presence of Invasive Exotic Grasses. Plant and Soil 281(1–2): 369–380. https://doi.org/10.1007/s11104-005-4826-3
- Hierro JL, Callaway RM (2003) Allelopathy and exotic plant invasion. Plant and Soil 256(1): 29–39. https://doi.org/10.1023/A:1026208327014
- Hu Y, Xu Z-W, Li M-Y, Croy JR, Zhang Z-Y, Li H-M, Guo W-H, Jiang X-L, Lu H-C, Guo X (2022) Increasing soil heterogeneity strengthens the inhibition of a native woody plant by an invasive congener. Plant and Soil 481(1–2): 677–690. https://doi.org/10.1007/s11104-022-05666-0
- Inderjit SD, Kaur H, Kalisz S, Bezemer TM (2021) Novel chemicals engender myriad invasion mechanisms. The New Phytologist 232(3): 1184–1200. https://doi.org/10.1111/nph.17685
- Inderjit SD, van der Putten WH (2010) Impacts of soil microbial communities on exotic plant invasions. Trends in Ecology and Evolution 25: 512–519. https://doi.org/10.1016/j.tree.2010.06.006
- Inderjit SD, Wardle DA, Karban R, Callaway RM (2011) The ecosystem and evolutionary contexts of allelopathy. Trends in Ecology and Evolution 26: 655–662. https://doi.org/10.1016/j.tree.2011.08.003
- Kardol P, Bezemer TM, van der Putten WH (2006) Temporal variation in plant-soil feedback controls succession. Ecology Letters 9(9): 1080–1088. https://doi.org/10.1111/j.1461-0248.2006.00953.x
- Li Y-P, Feng Y-L, Chen Y-J, Tian Y-H (2015) Soil microbes alleviate allelopathy of invasive plants. Science Bulletin 60(12): 1083–1091. https://doi.org/10.1007/s11434-015-0819-7
- Li S-P, Jia P, Fan S-Y, Wu Y, Liu X, Meng Y, Li Y, Shu W-S, Li J-T, Jiang L (2022) Functional traits explain the consistent resistance of biodiversity to plant invasion under nitrogen enrichment. Ecology Letters 25(4): 778–789. https://doi.org/10.1111/ele.13951
- Liu Y, Kleunen M, Cornelissen H (2017) Responses of common and rare aliens and natives to nutrient availability and fluctuations. Journal of Ecology 105(4): 1111–1122. https://doi.org/10.1111/1365-2745.12733
- Lorenzo P, Pereira CS, Rodríguez-Echeverría S (2013) Differential impact on soil microbes of allelopathic compounds released by the invasive Acacia dealbata Link. Soil Biology & Biochemistry 57: 156–163. https://doi.org/10.1016/j.soilbio.2012.08.018
- Ma J (2020) Alien invasive flora of China. Shanghai Jiao Tong University Press, Shanghai, China.
- McEwan RW, Arthur-Paratley LG, Rieske LK, Arthur MA (2010) A multi-assay comparison of seed germination inhibition by Lonicera maackii and co-occurring native shrubs. Flora – Morphology, Distribution. Flora (Jena) 205(7): 475–483. https://doi.org/10.1016/j.flora.2009.12.031
- Pellegrino E, Öpik M, Bonari E, Ercoli L (2015) Responses of wheat to arbuscular mycorrhizal fungi: A meta-analysis of field studies from 1975 to 2013. Soil Biology & Biochemistry 84: 210–217. https://doi.org/10.1016/j.soilbio.2015.02.020
- Qin TJ, Zhou J, Sun Y, Muller-Scharer H, Luo FL, Dong BC, Li HL, Yu FH (2020) Phylogenetic diversity is a better predictor of wetland community resistance to Alternanthera philoxeroides invasion than species richness. Plant Biology 22(4): 591–599. https://doi.org/10.1111/plb.13101
- Saia S, Jansa J (2022) Editorial: Arbuscular Mycorrhizal Fungi: The Bridge Between Plants, Soils, and Humans. Frontiers in Plant Science 13: 875958. https://doi.org/10.3389/fpls.2022.875958
- Sikes BA, Cottenie K, Klironomos JN (2009) Plant and fungal identity determines pathogen protection of plant roots by arbuscular mycorrhizas. Journal of Ecology 97(6): 1274–1280. https://doi.org/10.1111/j.1365-2745.2009.01557.x
- Vogelsang KM, Bever JD (2009) Mycorrhizal densities decline in association with nonnative plants and contribute to plant invasion. Ecology 90(2): 399–407. https://doi.org/10.1890/07-2144.1
- Wang C, Wei M, Wang S, Wu B, Cheng H (2020) Erigeron annuus (L.) Pers. and Solidago canadensis L. antagonistically affect community stability and community invasibility under the co-invasion condition. The Science of the Total Environment 716: 137128. https://doi.org/10.1016/j.scitotenv.2020.137128
- Wang J, Li SP, Ge Y, Wang XY, Gao S, Chen T, Yu FH (2022a) Darwin’s naturalization conundrum reconciled by changes of species interactions. Ecology 104(1): e3850. https://doi.org/10.1002/ecy.3850
- Wang Q, Li MY, Eller F, Luo YJ, Nong YL, Xing LJ, Xu ZW, Li HM, Lu HC, Guo X (2022b) Trait value and phenotypic integration contribute to the response of exotic Rhus typhina to heterogeneous nitrogen deposition: A comparison with native Rhus chinensis. The Science of the Total Environment 844: 157199. https://doi.org/10.1016/j.scitotenv.2022.157199
- Weremijewicz J, Sternberg LSLOR, Janos DP (2016) Common mycorrhizal networks amplify competition by preferential mineral nutrient allocation to large host plants. The New Phytologist 212(2): 461–471. https://doi.org/10.1111/nph.14041
- Xing L-J, Li M-Y, Jiang S-Y, Li W, Guo S-X, Li H-M, Guo X (2022) Comparison between the exotic Coreopsis grandiflora and native Dendranthema indicum across variable nitrogen deposition conditions. Acta Physiologiae Plantarum 44(8): 82. https://doi.org/10.1007/s11738-022-03417-3
- Xu Z, Zhong S, Yu Y, Wang Y, Cheng H, Du D, Wang C (2023) Rhus typhina L. triggered greater allelopathic effects than Koelreuteria paniculata Laxm under ammonium fertilization. Scientia Horticulturae 309: 111703. https://doi.org/10.1016/j.scienta.2022.111703
- Yuan L, Li J-M, Yu F-H, Oduor AMO, van Kleunen M (2021) Allelopathic and competitive interactions between native and alien plants. Biological Invasions 23(10): 3077–3090. https://doi.org/10.1007/s10530-021-02565-w
- Yuan L, Li J, van Kleunen M (2022) Competition induces negative conspecific allelopathic effects on seedling recruitment. Annals of Botany 130(6): 917–926. https://doi.org/10.1093/aob/mcac127
- Zhang S, Zhu W, Wang B, Tang J, Chen X (2011) Secondary metabolites from the invasive Solidago canadensis L. accumulation in soil and contribution to inhibition of soil pathogen Pythium ultimum. Applied Soil Ecology 48(3): 280–286. https://doi.org/10.1016/j.apsoil.2011.04.011
- Zhang F-J, Li Q, Chen F-X, Xu H-Y, Inderjit, Wan F-H (2017) Arbuscular mycorrhizal fungi facilitate growth and competitive ability of an exotic species Flaveria bidentis. Soil Biology & Biochemistry 115: 275–284. https://doi.org/10.1016/j.soilbio.2017.08.019
- Zhang P, Li B, Wu J, Hu S (2019) Invasive plants differentially affect soil biota through litter and rhizosphere pathways: A meta-analysis. Ecology Letters 22(1): 200–210. https://doi.org/10.1111/ele.13181
- Zhang Z, Liu Y, Yuan L, Weber E, van Kleunen M, Gurevitch J (2020) Effect of allelopathy on plant performance: A meta‐analysis. Ecology Letters 24(2): 348–362. https://doi.org/10.1111/ele.13627
Supplementary material
Responses of the aboveground biomass of native plant species to the allelopathy (n = 10)
Data type: tif
Explanation note: The boxes represent the range of maximum and minimum values, with a bold horizontal line indicating the average value. The plant species include Chenopodium album L. (A), Vitex negundo L. (B), Rhus chinensis Mill. (C), and Acer truncatum Bunge (D). There were five different concentrations of the root aqueous extracts of R. typhina including 0 g L− 1 (CK), 10 g− 1 (LRE), 20 g L− 1 (NRE), 30 g L− 1 (MRE), and 40 g L– 1 (HRE). The asterisk (*) indicates a significant difference (p ≤ 0.05) with Duncan’s test.