





(C) 2011 Petr Pyšek. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
This issue has been dealt with in detail in some invasion biology literature from other regions of the world, a fact that is not acknowledged in
The above indicates that the problem may be more political/management related, rather than scientific, i.e. rather related to large databases covering the whole country and built for pratical purposes, such as USDA and NRCS, referred to by
European data (based on the DAISIE project;
On the continent as a whole there are 3749 naturalized (sensu
At the scale of individual countries, for example considering incorrectly only those aliens with their region of origin entirely outside of Europe would move 522 of the neophytes (alien species introduced since the beginning of the 16th century) in the Czech Republic (49.9% of the total number of neophytes) into the “native” category. This would reduce the number of neophytes recorded from 1046 to 524, and their proportional contribution to the country‘s entire flora from 25.3% to 12.7% (based on data in
The problem is essentially one of scale. There is no specific scale at which invasions should be studied, and the patterns and processes found by researchers differ according to the grain of the study (e.g.,
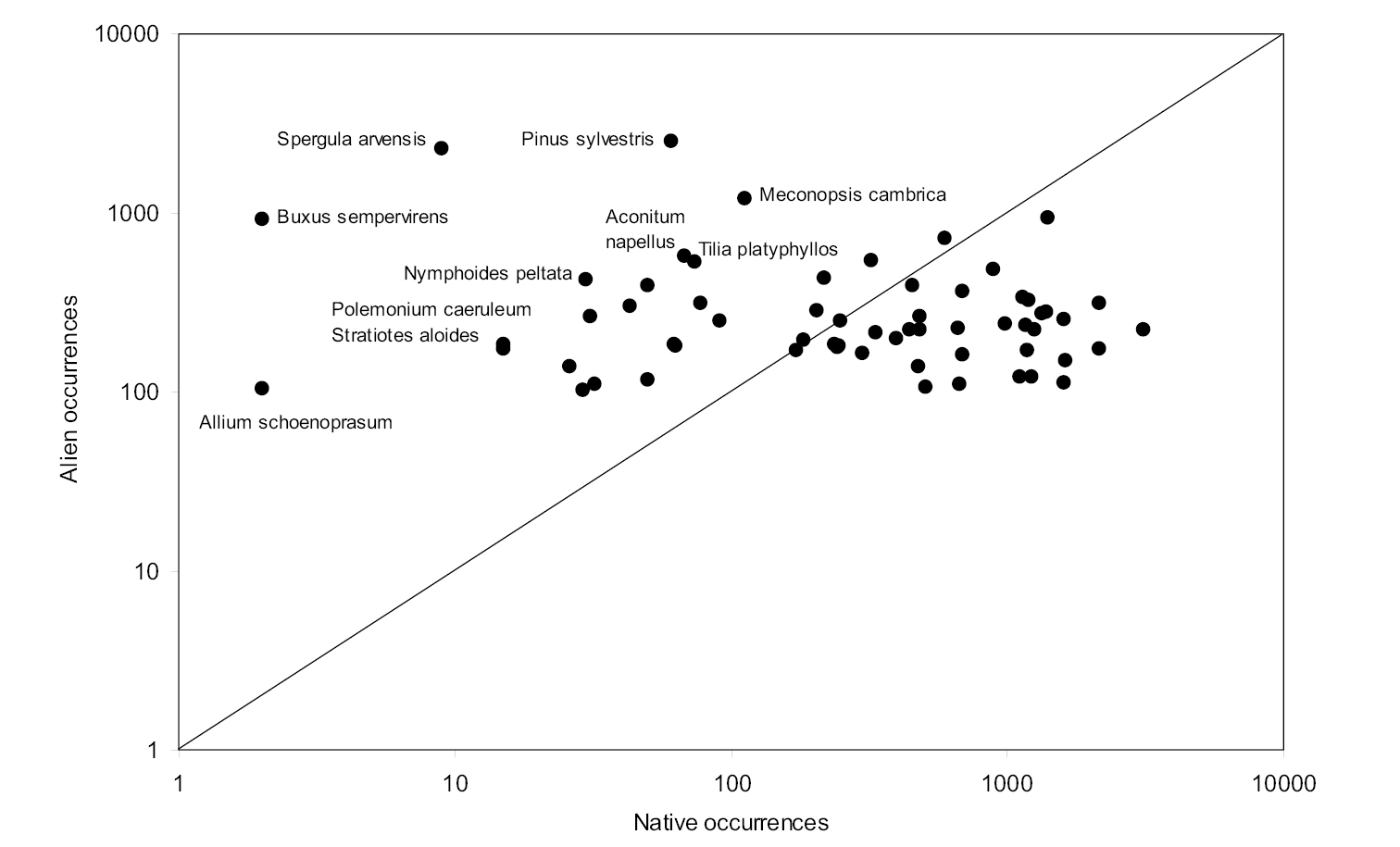
Number of 10 ×10 km grid cells in which species native to the British Isles, that also occur in localities where their occurrence is considered alien, were mapped in 1987–1999. Species above and below the unity line are recorded more frequently as “aliens” and “natives”, respectively. Based on data from
As pointed by
In terms of science, lumping both groups of origin together may obscure ecological analyses since the groups differ in many respects; the data gathered for Europe again allow for a more detailed insight. Besides the fact that biased species richness complicates comparisons between regions for which the data was collected by one approach or the other (
Moreover, comparisons seeking for determinants of regional invasibility based on floras of whole states or groups of states can be misleading. This is because high environmental heterogeneity across a large area may hide the patterns that could be seen if comparing alien species richness within smaller, more homogeneous areas that are more suitable for revealing causal connections between environmental factors, anthropic disturbance, history, and numbers of alien species (
In invasion biology, it is becoming increasingly important to synthesize existing data on alien species abundances and distributions (
The author was supported by long-term institutional research plans no. AV0Z60050516 (Academy of Sciences of the Czech Republic), no. MSM0021620828 (Ministry of Education of the Czech Republic), and by the Praemium Academiae award from the Academy of Sciences of the Czech Republic.