Research Article |
|
Corresponding author: Marek Šmejkal ( marek.smejkal@hbu.cas.cz ) Academic editor: Ingolf Kühn
© 2024 Marek Šmejkal, Kiran Thomas, Vladimír Kořen, Jan Kubečka.
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Šmejkal M, Thomas K, Kořen V, Kubečka J (2024) The 50-year history of anglers' record catches of genus Carassius: circumstantial evidence of wiping out the native species by invasive conspecific. NeoBiota 92: 111-128. https://doi.org/10.3897/neobiota.92.121288
|
Abstract
Successful invasive non-native fish species can cause enormous damage to native biodiversity. In mainland Europe, the introduction of the gibel carp (Carassius gibelio) has led to a decline in populations of the formerly widespread native crucian carp (C. carassius). Both invasive and native species develop two phenotypes, namely stunted and deep-bodied, which depend on the intensity of competition and predation in the water body. The deep-bodied phenotype is associated with a more diverse fish community composition, can attain large sizes and is very attractive to recreational anglers. This study analysed trends in the record sizes of native crucian carp and invasive gibel carp (individuals close to the maximum attainable size of the species) reported by recreational anglers over the last 50 years in Czechia, recording the invasion of gibel carp from its beginnings to the fully established population phase. The study provides circumstantial evidence that gibel carp is behind transition from the relative abundance of large crucian carp to near extirpation, while large gibel carp have taken over the reports of record catches in the genus Carassius. This indicates that the crucian carp, which is currently classified as critically endangered in the Red List of Czechia, has very limited possibilities to realise its deep-bodied phenotype. It also shows the potential of using data from recreational anglers for mapping invasion processes and as a source of relatively localised information on endangered species.
Graphical abstract
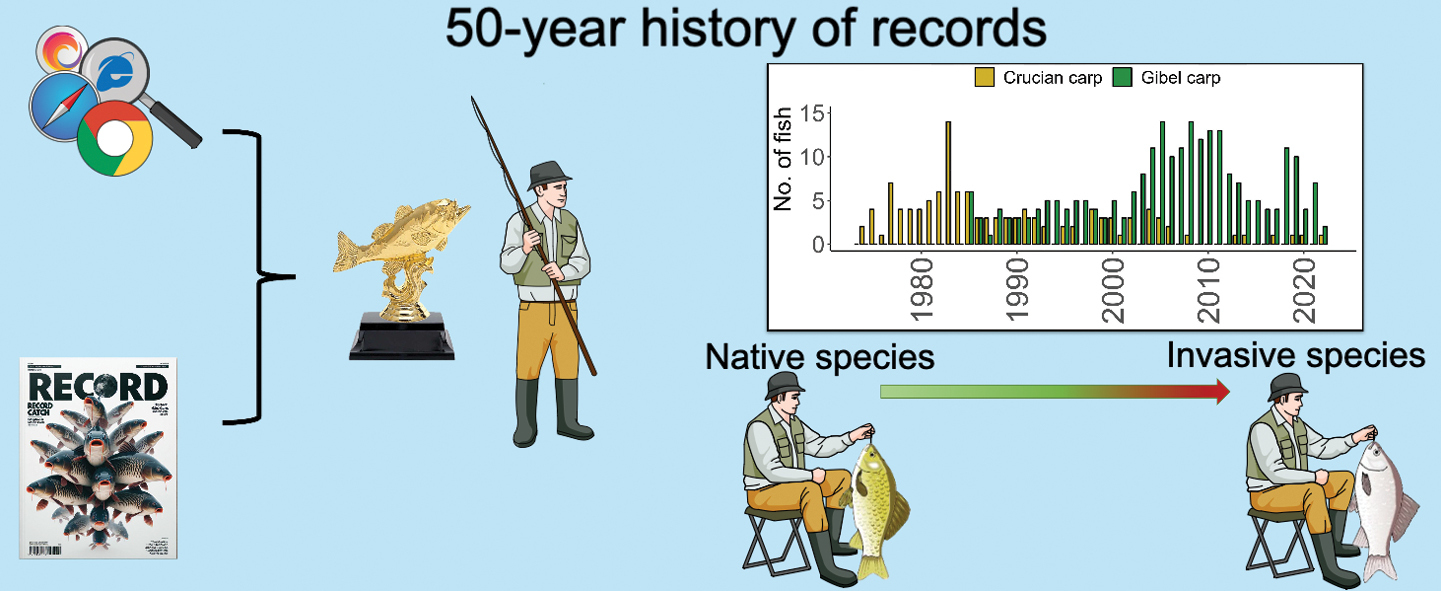
Key words
Aquatic conservation, culturomics, crucian carp, gibel carp, iEcology, Prussian carp
Introduction
Invasive non-native species can have an enormous impact on freshwater ecosystems, displacing native species or even causing their complete extinction (
Fish of the Cyprinidae family were the first to be spread outside their native waters (
Due to limited resources in monitoring the progress of invasive non-native species, it has proven useful to utilise knowledge, photos and video recordings collected by people through approaches such as citizen science, culturomics and iEcology (
Despite the great attention paid by recreational anglers to large species and individuals (
The crucian carp used to be one of the most abundant species in small European lentic waters; however, it declined due to habitat reduction and the invasion of the gibel carp in Western and Central Europe (
This study thus attempts to retrospectively map the disappearance of the large-sized crucian carp after the invasion of the gibel carp, which was not well documented by the regular monitoring activities of scientists and nature conservation authorities. To this end, recreational angling magazines and websites dedicated to record catches were scrutinised for focal species. In addition to information on fish and catches, recreational anglers also provided photos of their record catches. The study selected evidence of record catches of native crucian carp and invasive gibel carp and used common bream (Abramis brama) and roach (Rutilus rutilus) as a reference dataset. These species were not as severely affected by the invasion of the gibel carp in Czechia as native species in southern latitudes and the gibel carp did not cause a significant decline in the populations of these species in reservoirs and large rivers (
Materials and methods
Record catches extraction and verification
The first dataset compiled for the study includes reported catch records of the focal species for which the recreational angling magazines “Rybářství”, “Rybář”, “Český Rybář”, “Sportovní Rybářství”, “Kajman” and the recreational angler´s website “mrk.cz” were examined for evidence of record fish of the species crucian carp, gibel carp, common bream and roach from the years 1973–2022, resulting in a dataset spanning 50 years. The submitted photos, together with the total length (TL, in cm), weight (kg; W) and angling area code, were first reviewed by a magazine editor-in-chief or an expert (an experienced angler and often an authority from the Czech or Moravian Anglers Union, or from the anglers’ community on the mrk.cz website) before being officially admitted to the competition. Selected photos appeared directly in the magazines, while the rest of the record catches were not selected for publication in the magazines and were included in the annual summarised statistics of record catches. Both datasets were extracted in a standardised manner, focusing on the presence of TL, W, species, water type (lentic, lotic) and numerical angling area identifier (code unique to a particular water body or river section, if available in the record). In addition, due to the similarity between crucian carp and gibel carp, the authors’ team reviewed all available photos to assess the reliability of the dataset and changed the category from crucian carp to gibel carp (or vice versa) where appropriate. We also extracted the central GPS position of the angling site and the total area in ha and assigned the angling ground to three existing watersheds in Czechia (Elbe, Danube and Odra; Suppl. material
The angling sites are part of the Czech and Moravian Anglers Unions, which are large organisations with around 330,000 members and whose angling grounds cover the catchment areas of the Elbe, Danube (Morava) and Odra Rivers in Czechia. There are 76,000 kilometres of watercourses and 107 reservoirs in Czechia. The area of waters totals 42,000 hectares, both in lotic and lentic ecosystems and is accessible for recreational angling with a licence.
Statistical analyses
To estimate whether the number of record-sized individuals of a given species follows any pattern in the time series, the data were tested with the funtimes package (
In addition, the records of native crucian carp and invasive gibel carp were analysed to determine whether they differ in their maximum recorded length and weight. The crucian carp and gibel carp datasets were tested for normality assumptions using the Shapiro-Wilk normality test. As the data were not normally distributed, the Mann-Whitney U-test was used for all data regardless of the year of capture.
Generalised additive models (GAM) were used to assess trends in fish size during the study period (
Generalised additive models were used to plot the points of records of all four species for each decade, using the number of reported catches in each period as the response variable and their GPS locations as the explanatory variable, by creating contour plots (
Results
In total, the dataset contained 982 records in the period 1973–2022, with 124 records of native crucian carp (mean TL = 39.8 ± 4.3 cm, mean W = 1.46 ± 0.43 kg), 248 invasive gibel carp (TL = 44.5 ± 4.0 cm; W = 1.82 ± 0.50 kg), 369 common bream (TL = 62.3 ± 5.7 cm, W = 3.29 ± 0.87 kg) and 241 roach (TL = 40.8 ± 3.5 cm, W = 1.09 ± 0.27 kg). Of the total number of catches, 66.9% of the native crucian carp were caught in lentic waters, while the figures for invasive gibel carp, common bream and roach were 62.9%, 42.5% and 36.5%, respectively. The highest contribution to the dataset was made by the magazine “Rybářství”(700), followed by Kajman (103), mrk.cz (93), Český Rybář (48), Rybář (34) and Sportovní Rybářství (4). When checking the available photos of crucian carp and gibel carp, 27 and 98 photos were obtained, respectively. The reliability of species identification on these photos reached 63% for crucian carp and 100% for gibel carp. All misidentifications were made after 1993, while all 10 crucian carp records with photos were confirmed as crucian carp before that year.
Trend analyses of reported record fishes
The test for any trend on all four species indicated that all species contain a significant trend in their data (WAVK test: crucian carp p < 0.001; gibel carp p < 0.001; common bream < 0.001; roach < 0.05). Trends in number of record crucian carp reported by recreational anglers declined sharply after 2005 and, for these data and the best fit, we used a model with moving window (MW) 7 and a polynomial fit of degree 11 (WAVK test = 34.098, p < 0.001). Reports of invasive gibel carp first appeared in 1985 and were comparable in number to native crucian carp between 1985 and 2000. Since then, reports of invasive gibel carp have become very dominant in terms of record sizes (Fig.

The number of record catches extracted from angling magazines and websites between 1973 and 2022 for A the native crucian carp (Carassius carassius), invasive gibel carp (C. gibelio) and reference fish species B common bream (Abramis brama) and C roach (Rutilus rutilus). The period captures the invasion phase of gibel carp from early invasion phase to fully established, as well as the current impact on the native and critically endangered crucian carp in Czechia.
Trends in species maximum sizes
The reported native crucian carp were, on average, smaller than invasive gibel carp in both length (39.8 ± 4.3 cm vs. 44.5 ± 4.0 cm, W = 6167.0, p < 0.001; Fig.
The parametric coefficient of general additive models for record catches of native crucian carp (Carassius carassius), invasive gibel carp (C. gibelio), common bream (Abramis brama) and roach (Rutilus rutilus) with response variable of total length (upper table) and weight (lower table). The significance of smooth term on variable year was < 0.001 for both models and explained deviance was 83.4 and 71.8%, respectively.
| Estimate | t-value | p-value | |
| GAM Length | |||
| Intercept | 40.174 | 74.966 | <0.001 |
| Species:Roach | 0.383 | 0.709 | NS |
| Species:Bream | 22.069 | 43.021 | <0.001 |
| Species:Gibel | 3.812 | 6.876 | <0.001 |
| Angling ground size | 0.001 | 2.369 | 0.0181 |
| Type: Lotic | -0.088 | -0.267 | NS |
| Basin: Elbe | 0.298 | 0.840 | NS |
| Basin: Odra | -0.820 | -1.152 | NS |
| GAM Weight | |||
| Intercept | 1.408 | 19.973 | <0.001 |
| Species:Roach | -0.420 | -5.908 | <0.001 |
| Species:Bream | 1.810 | 26.838 | <0.001 |
| Species:Gibel | 0.314 | 4.305 | <0.001 |
| Angling ground size | 7.094e-05 | 2.157 | 0.031 |
| Type: Lotic | 0.001 | 0.198 | NS |
| Basin: Elbe | 0.107 | 2.280 | 0.029 |
| Basin: Odra | -0.041 | -0.440 | NS |

The comparison of reported record catches size in the 50-year dataset. The maximum attainable size of the invasive gibel carp (Carassius gibelio) significantly exceeds the maximum size of the native crucian carp (C. carassius) in Czechia in both A total length and B weight. The boxplot boundaries represent upper and lower quartiles; the thick lines represent medians and the whiskers represent 1.5 times the interquartile range. Violin plots represent kernel density distribution.

The GAM trend-line of record catches size (total length) in the 50-year dataset. The GAM fit has been computed with the stat_smooth function. The data suggest: A unimodal response in the native crucian carp (Carassius carassius) with time B increase maximum attainable size in the invasive gibel carp (C. gibelio) C stabilised size limits in common bream (Abramis brama) and D maximum in 1990 in roach (Rutilus rutilus).

The GAM trend-line of record catches size (weight) in the 50-year dataset. The fit has been computed with the smooth function with a value of k = 1 to avoid overfitting. The data suggest A unimodal response in the native crucian carp (Carassius carassius) with time B moderate increase in maximum attainable size in the invasive gibel carp (C. gibelio) C relatively stabilised size limits in common bream (Abramis brama) and D fluctuating trend in roach (Rutilus rutilus).
The GAM contour plots showed the concentration of large-sized fish mainly in the area of the lowland rivers and the interconnected oxbows. While common bream and gibel carp were almost ubiquitous, record-sized crucian carp and roach showed a more aggregated distribution with few hotspots in Czechia (Fig.

Contour plots based on GAMs for A native crucian carp (Carassius carassius) B invasive gibel carp (C. gibelio) C common bream (Abramis brama) and D roach (Rutilus rutilus) for all study period, where the number of reported record catches in a given area was used as the response variable and their GPS locations as the explanatory variable. Each line (or contour) represents the reported catches in that region within the country. The proximity of the lines indicates the steepness of the gradient. Colours show the abundance of records with red indicating the highest abundance.
Discussion
Given the speed at which invasions are progressing in the aquatic environment, it appears that utilising the information collected from citizens can help combat the problem (
Trends in occurrence and size with relation to species ecology
Both native crucian carp and invasive gibel carp were found more frequently in the lentic waters, which is consistent with their ecology (
While the records of both reference species showed a relatively stable trend around the same average value, both native crucian carp and invasive gibel carp showed GAM trend-lines indicating changes in maximum size. For crucian carp, the unimodal response with a decline in recorded maximum size in recent years suggests either growth limitations or possible confusion with invasive gibel carp or hybrids between crucian carp and gibel carp around 2000. Both options are possible, as the growth restriction may be caused either by increased interspecific competition due to the invasion of the gibel carp (
Hybridisation is also a likely explanation, as both species form hybrids under certain circumstances (
The sequence of gibel carp invasion in Czechia
The gibel carp was accidentally introduced to Hungary together with the commercial Asian fish species (Hypophthalmichthys molitrix, H. nobilis, Ctenopharyngodon idella) in the 1950s (
There are currently three invasive species of the genus Carassius living in Czechia, the goldfish (including feral form), C. langsdorfii in addition to already mentioned gibel carp (
Engaging public in conservation of crucian carp
The passion for enhancing native crucian carp populations was first brought to public attention in England, where Peter Rolfe launched his attempt as a pilot angler to reintroduce the species to Norfolk waters and promote the existence of exceptionally large fish in ponds (
Although the conservation and fisheries management described above is artificial, it has its roots in the life-history strategy of the crucian carp. In the floodplain rivers, the population has a metapopulation structure with a rare deep-bodied phenotype in multi-species community of large water bodies or lowland rivers (
Culturomics role in fish conservation with emphasis on large-size fish
Culturomics in conservation has been shown to be an effective means of raising public understanding, framing conservation issues and engaging people in timely environmental monitoring (
Acknowledgements
We sincerely thank all those people reporting their record catches to the magazine and, thus, helping to capture the decline of large-sized crucian carp. We are especially grateful to Tomáš Matýsek, who, with his passion to record fish statistics, helped to reveal the patterns captured in the data.
Additional information
Conflict of interest
The authors have declared that no competing interests exist.
Ethical statement
This study did not involve handling of animals or personal information.
Funding
The collection of data resulting in this manuscript has been supported by the programme of Regional Cooperation of the Czech Academy of Sciences (R200962201) and the Research Programme Strategy AV21 Water for life for valuable support.
Author contributions
Marek Šmejkal: Conceptualisation, Investigation, Data curation, Formal analysis, Writing Original draft, Writing, Review & Editing, Visualisation. Kiran Thomas: Formal analysis, Visualisation, Writing, Review & Editing. Vladimír Kořen: Conceptualisation, Writing, Review & Editing. Jan Kubečka: Conceptualisation, Data curation, Writing, Review & Editing.
Author ORCIDs
Marek Šmejkal https://orcid.org/0000-0002-7887-6411
Data availability
Data will be shared by authors upon reasonable request.
References
- Almena IS, Balzani P, Carneiro L, Cuthbert RN, Macedo R, Tarkan AS, Ahmed DA, Bang A, Bacela-Spychalska K, Bailey SA, Baudry T, Ballesteros L, Bortolus A, Briski E, Britton JR, Buric M, Camacho-Cervantes M, Cano-Barbacil C, Copilaș-Ciocianu D, Coughlan N, Courtois P, Csabai Z, Dalu T, De Santis V, Dickey JWE, Dimarco R, Falk-Andersson J, Fernandez R, Florencio M, Franco ACS, Garcia-Berthou E, Giannetto D, Glavendekic M, Grabowski M, Heringer G, Herrera I, Wei H, Kamelamela KL, Kirichenko NI, Kouba A, Kourantidou M, Kurtul I, Laufer G, Lipták B, Liu C, Lopez-López E, Lozano V, Mammola S, Marchini A, Meshkova V, Meyerson L, Milardi M, Musolin DL, Nuñez M, Oficialdegui FJ, Patoka J, Pattision Z, Petrusek A, Pincheira-Donoso D, Piria M, Probert A, Rasmussen JJ, Renault D, Ribeiro F, Rilov G, Robinson TB, Sanchez A, Schwindt E, South J, Stoett P, Verreycken H, Vilizzi L, Wang Y-J, Watari Y, Wehi PM, Weiperth A, Wiberg-Larsen P, yapici S, Yoğurtçuoğlu B, Zenni R, Galil BS, Dick JTA, Russell J, Ricciardi A, Simberloff D, Bradshaw CJA, Haubrock PJ (2023) Taming the terminological tempest in invasion science. EcoEvoRxiv 17: 1–83. https://doi.org/10.32942/X24C79
- Arlinghaus R, Tillner R, Bork M (2015) Explaining participation rates in recreational fishing across industrialised countries. Fisheries Management and Ecology 22(1): 45–55. https://doi.org/10.1111/fme.12075
- Arlinghaus R, Abbott JK, Fenichel EP, Carpenter SR, Hunt LM, Alós J, Klefoth T, Cooke SJ, Hilborn R, Jensen OP, Wilberg MJ, Post JR, Manfredo MJ (2019) Governing the recreational dimension of global fisheries. Proceedings of the National Academy of Sciences of the United States of America 116(12): 5209–5213. https://doi.org/10.1073/pnas.1902796116
- Augustin NH, Sauleau EA, Wood SN (2012) On quantile quantile plots for generalized linear models. Computational Statistics & Data Analysis 56(8): 2404–2409. https://doi.org/10.1016/j.csda.2012.01.026
- Auwerx J, Van Wichelen J, De Charleroy D (2021) Prussian carp beats crucian carp in the struggle for existence. In: 9th EPCN Conference, London 2021.
- Balon EK (2004) About the oldest domesticates among fishes. Journal of Fish Biology 65(s1): 1–27. https://doi.org/10.1111/j.0022-1112.2004.00563.x
- Bartoň D, Šmejkal M (2022) Zhodnocení stavu reofilních ryb v povodí Vltavy a klíčových faktorů ovlivňující jejich populace [Evaluation of rheophilic fish status in Vltava catchment and key factors influencing their populations - Report]. Výzkumná zpráva pro státní podnik Povodí Vlt. České Budějovice.
- Beardmore B, Hunt LM, Haider W, Dorow M, Arlinghaus R (2015) Effectively managing angler satisfaction in recreational fisheries requires understanding the fish species and the anglers. Canadian Journal of Fisheries and Aquatic Sciences 72(4): 500–513. https://doi.org/10.1139/cjfas-2014-0177
- Birdsong M, Hunt LM, Arlinghaus R (2021) Recreational angler satisfaction: What drives it? Fish and Fisheries 22(4): 682–706. https://doi.org/10.1111/faf.12545
- Blackburn TM, Pyšek P, Bacher S, Carlton JT, Duncan RP, Jarošík V, Wilson JRU, Richardson DM (2011) A proposed unified framework for biological invasions. Trends in Ecology & Evolution 26(7): 333–339. https://doi.org/10.1016/j.tree.2011.03.023
- Blažka P (1958) The anaerobic metabolism of fish. Physiological Zoology 31(2): 117–128. https://doi.org/10.1086/physzool.31.2.30155385
- Boon JS, Vaudin G, Millward-Hopkins H, O’Leary BC, McClean CJ, Stewart BD (2024) Shifts in the size and distribution of marine trophy fishing world records. Aquatic Conservation 34(1): e4051. https://doi.org/10.1002/aqc.4051
- Boukal DS, Jankovský M, Kubečka J, Heino M (2012) Stock–catch analysis of carp recreational fisheries in Czech reservoirs: Insights into fish survival, water body productivity and impact of extreme events. Fisheries Research 119–120: 23–32. https://doi.org/10.1016/j.fishres.2011.12.003
- Brönmark C, Miner JG (1992) Predator-induced phenotypical change in body morphology in crucian carp. Science 258(5086): 1348–1350. https://doi.org/10.1126/science.258.5086.1348
- Brönmark C, Paszkowski CA, Tonn WM, Hargeby A (1995) Predation as a determinant of size structure in populations of crucian carp (Carassius carassius) and tench (Tinca tinca). Ecology Freshwater Fish 4(2): 85–92. https://doi.org/10.1111/j.1600-0633.1995.tb00121.x
- Busst GMA, Britton JR (2015) Quantifying the growth consequences for crucian carp Carassius carassius of competition from non-native fishes. Ecology Freshwater Fish 24(3): 489–492. https://doi.org/10.1111/eff.12155
- Busst GMA, Britton JR (2017) Comparative trophic impacts of two globally invasive cyprinid fishes reveal species-specific invasion consequences for a threatened native fish. Freshwater Biology 62(9): 1587–1595. https://doi.org/10.1111/fwb.12970
- Cooke SJ, Suski CD (2005) Do we need species-specific guidelines for catch-and-release recreational angling to effectively conserve diverse fishery resources? Biodiversity and Conservation 14(5): 1195–1209. https://doi.org/10.1007/s10531-004-7845-0
- Copp G, Sayer CD (2010) Norfolk Biodiversity Action Plan – Local species action plan for crucian carp (Carassius carassius).
- Copp G, Tarkan S, Godard M, Edmonds N, Wesley K (2010) Preliminary assessment of feral goldfish impacts on ponds, with particular reference to native crucian carp. Aquatic Invasions 5(4): 413–422. https://doi.org/10.3391/ai.2010.5.4.11
- de Meo I, Østbye K, Kahilainen KK, Hayden B, Fagertun CHH, Poléo ABS (2021) Predator community and resource use jointly modulate the inducible defense response in body height of crucian carp. Ecology and Evolution 11(5): 2072–2085. https://doi.org/10.1002/ece3.7176
- de Meo I, Østbye K, Kahilainen KK, Hayden B, Magnus M, Poléo ABS (2022) Resource use of crucian carp along a lake productivity gradient is related to body size, predation risk, and resource competition. Ecology Freshwater Fish 32(1): 10–22. https://doi.org/10.1111/eff.12668
- Emmrich M, Pédron S, Brucet S, Winfield IJ, Jeppesen E, Volta P, Argillier C, Lauridsen TL, Holmgren K, Hesthagen T, Mehner T (2014) Geographical patterns in the body-size structure of European lake fish assemblages along abiotic and biotic gradients. Journal of Biogeography 41(12): 2221–2233. https://doi.org/10.1111/jbi.12366
- Fedorčák J, Križek P, Koščo J (2023) Which factors influence spatio–temporal changes in the distribution of invasive and native species of genus Carassius? Aquatic Invasions 18(2): 219–230. https://doi.org/10.3391/ai.2023.18.2.105240
- García-Asorey MI, Escati-Peñaloza G, Parma AM, Pascual MA, Marshall CT (2011) Conflicting objectives in trophy trout recreational fisheries: Evaluating trade-offs using an individual-based model. Canadian Journal of Fisheries and Aquatic Sciences 68(11): 1892–1904. https://doi.org/10.1139/f2011-108
- Gozlan RE, St.-Hilaire S, Feist SW, Martin P, Kent ML (2005) Disease threat to European fish. Nature 435(7045): 1046–1046. https://doi.org/10.1038/4351046a
- Grabowska J, Błońska D, Kati S, Nagy SA, Kakareko T, Kobak J, Antal L (2019) Competitive interactions for food resources between the invasive Amur sleeper (Perccottus glenii) and threatened European mudminnow (Umbra krameri). Aquatic Conservation 29(12): 2231–2239. https://doi.org/10.1002/aqc.3219
- Gurevitch J, Padilla DK (2004) Are invasive species a major cause of extinctions? Trends in Ecology & Evolution 19(9): 470–474. https://doi.org/10.1016/j.tree.2004.07.005
- Halačka K, Lusková V, Lusk S (2003) Carassius gibelio in fish communities of the Czech Republic. Ecohydrology & Hydrobiology 3: 133–138.
- Hensel K (1971) Some notes on the systematic status of Carassius auratus gibelio (Bloch, 1782) with further record of this fish from the Danube River in Czechoslovakia. Věstník Československé Společnosti Zoologické 3: 186–198.
- Holčík J (1980) Possible Reason for the Expansion of Carassius auratus (Linnaeus, 1758) (Teleostei, Cyprinidae) in the Danube River Basin. Internationale Revue der Gesamten Hydrobiologie 65(5): 673–679. https://doi.org/10.1002/iroh.19800650507
- Holopainen IJ, Hyvärinen H (1985) Ecology and physiology of crucian carp [Carassius carassius (L.)] in small Finnish ponds with anoxic conditions in winter. SIL Proceedings 22(4): 2566–2570. https://doi.org/10.1080/03680770.1983.11897726
- Holopainen IJ, Tonn WM, Paszkowski CA (1997) Tales of two fish: The dichotomous biology of crucian carp (Carassius carassius (L.)) in northern Europe. Annales Zoologici Fennici 34: 1–22.
- Hutt CP, Hunt KM, Anderson DK (2013) Measurement of angler catch-related attitudes: An assessment of model structure and metric invariance. Leisure Sciences 35(4): 382–398. https://doi.org/10.1080/01490400.2013.797716
- Jarić I, Correia RA, Brook BW, Buettel JC, Courchamp F, Di Minin E, Firth JA, Gaston KJ, Jepson P, Kalinkat G, Ladle R, Soriano-Redondo A, Souza AT, Roll U (2020a) iEcology: Harnessing Large Online Resources to Generate Ecological Insights. Trends in Ecology & Evolution 35(7): 630–639. https://doi.org/10.1016/j.tree.2020.03.003
- Jarić I, Roll U, Arlinghaus R, Belmaker J, Chen Y, China V, Douda K, Essl F, Jähnig SC, Jeschke JM, Kalinkat G, Kalous L, Ladle R, Lennox RJ, Rosa R, Sbragaglia V, Sherren K, Šmejkal M, Soriano-Redondo A, Souza AT, Wolter C, Correia RA (2020b) Expanding conservation culturomics and iEcology from terrestrial to aquatic realms. PLoS Biology 18(10): e3000935. https://doi.org/10.1371/journal.pbio.3000935
- Jarić I, Bellard C, Correia RA, Courchamp F, Douda K, Essl F, Jeschke JM, Kalinkat G, Kalous L, Lennox RJ, Novoa A, Proulx R, Pyšek P, Soriano–Redondo A, Souza AT, Vardi R, Veríssimo D, Roll U (2021) Invasion Culturomics and iEcology. Conservation Biology 35(2): 447–451. https://doi.org/10.1111/cobi.13707
- Kahle D, Wickham H (2013) ggmap: Spatial visualization with ggplot2. The R Journal 5(1): 144–161. https://doi.org/10.32614/RJ-2013-014
- Kalous L, Šlechtová Jr V, Bohlen J, Petrtýl M, Švátora M (2007) First European record of Carassius langsdorfii from the Elbe basin. Journal of Fish Biology 70(sa): 132–138. https://doi.org/10.1111/j.1095-8649.2006.01290.x
- Kalous L, Bohlen J, Rylková K, Petrtý M (2012) Hidden diversity within the Prussian carp and designation of a neotype for Carassius gibelio (Teleostei: Cyprinidae). Ichthyological Exploration of Freshwaters 23: 11–18.
- Kalous L, Rylková K, Bohlen J, Šanda R, Petrtýl M (2013) New mtDNA data reveal a wide distribution of the Japanese ginbuna Carassius langsdorfii in Europe. Journal of Fish Biology 82(2): 703–707. https://doi.org/10.1111/j.1095-8649.2012.03492.x
- Keszte S, Ferincz A, Tóth-Ihász K, Balogh RE, Staszny Á, Hegyi Á, Takács P, Urbanyi B, Kovács B (2021) Mitochondrial sequence diversity reveals the hybrid origin of invasive gibel carp (Carassius gibelio) populations in Hungary. PeerJ 9: e12441. https://doi.org/10.7717/peerj.12441
- Knytl M, Kalous L, Rylková K, Choleva L, Merila J, Ráb P (2018) Morphologically indistinguishable hybrid Carassius female with 156 chromosomes: A threat for the threatened crucian carp, C. carassius, L. PLoS ONE 13(1): e0190924. https://doi.org/10.1371/journal.pone.0190924
- Knytl M, Forsythe A, Kalous L (2022) A fish of multiple faces, which show us enigmatic and incredible phenomena in nature: Biology and cytogenetics of the genus Carassius. International Journal of Molecular Sciences 23(15): 8095. https://doi.org/10.3390/ijms23158095
- Kubečka J (1989) The spreading of the german carp, Carassius auratus (Linnaeus) in the middle Elbe River. Muzeum a současnost, ser. natur. 3: 43–50.
- Ladle RJ, Correia RA, Do Y, Joo G-J, Malhado AC, Proulx R, Roberge J-M, Jepson P (2016) Conservation culturomics. Frontiers in Ecology and the Environment 14(5): 269–275. https://doi.org/10.1002/fee.1260
- Locker A (2014) The social history of coarse angling in England AD 1750–1950. Anthropozoologica 49(1): 99–107. https://doi.org/10.5252/az2014n1a07
- Löki V, Nagy J, Neményi Z, Hagyó A, Nagy A, Vitál Z, Mozsár A, Lukács BA (2023) Exploring ecological knowledge in recreational fishing for conservation purposes: A literature review. Global Ecology and Conservation 48: e02697. https://doi.org/10.1016/j.gecco.2023.e02697
- Lusk S, Lusková V, Hanel L (2010) Alien fish species in the Czech Republic and their impact on the native fish fauna. Folia Zoologica 59(1): 57–72. https://doi.org/10.25225/fozo.v59.i1.a9.2010
- Lyach R (2022) Increasing dominance of non-native fishes in the yield of central European streams and rivers. Fisheries Research 254: 106433. https://doi.org/10.1016/j.fishres.2022.106433
- Lyubchich V, Gel YR (2023) funtimes: Functions for Time Series Analysis. R package version 9.1.
- Lyubchich V, Gel YR, El-Shaarawi A (2013) On detecting non-monotonic trends in environmental time series: A fusion of local regression and bootstrap. Environmetrics 24(4): 209–226. https://doi.org/10.1002/env.2212
- Musil J, Jurajda P, Adámek Z, Horký P, Slavík O (2010) Non-native fish introductions in the Czech Republic - species inventory, facts and future perspectives. Journal of Applied Ichthyology 26: 38–45. https://doi.org/10.1111/j.1439-0426.2010.01500.x
- Naylor RL, Williams SL, Strong DR (2001) Aquaculture-A Gateway for exotic species. Science 294(5547): 1655–1656. https://doi.org/10.1126/science.1064875
- Papoušek I, Vetešník L, Halačka K, Lusková V, Humpl M, Mendel J (2008) Identification of natural hybrids of gibel carp Carassius auratus gibelio (Bloch) and crucian carp Carassius carassius (L.) from lower Dyje River floodplain (Czech Republic). Journal of Fish Biology 72(5): 1230–1235. https://doi.org/10.1111/j.1095-8649.2007.01783.x
- Pentyliuk N, Schmidt B, Poesch MS, Green SJ (2023) Recreational angler reporting as a tool for tracking the distribution of invasive Prussian carp (Carassius gibelio). Conservation Science and Practice 5(1): e12850. https://doi.org/10.1111/csp2.12850
- Perdikaris C, Ergolavou A, Gouva E, Nathanailides C, Chantzaropoulos A, Paschos I (2012) Carassius gibelio in Greece: The dominant naturalised invader of freshwaters. Reviews in Fish Biology and Fisheries 22(1): 17–27. https://doi.org/10.1007/s11160-011-9216-8
- Pettersson LB, Bronmark C (1997) Density-dependent costs of an inducible morphological defense in crucian carp. Ecology 78(6): 1805–1815. https://doi.org/10.1890/0012-9658(1997)078[1805:DDCOAI]2.0.CO;2
- Piironen J, Holopainen IJ (1986) A note on seasonality in anoxia tolerance of crucian carp (Carassius carassius (L.)) in the laboratory. Annales Zoologici Fennici 23: 335–338. https://doi.org/10.2307/23736015
- Pinder AC, Raghavan R, Britton JR (2015) Efficacy of angler catch data as a population and conservation monitoring tool for the flagship Mahseer fishes (Tor spp.) of Southern India. Aquatic Conservation 25(6): 829–838. https://doi.org/10.1002/aqc.2543
- R Core Team (2023) R: A Language and Environmental for Statistical Computing R Development Core Team, R: a language and environment for statistical computing.
- Ribeiro F, Rylková K, Moreno-Valcárcel R, Carrapato C, Kalous L (2015) Prussian carp Carassius gibelio: A silent invader arriving to the Iberian Peninsula. Aquatic Ecology 49(1): 99–104. https://doi.org/10.1007/s10452-015-9508-5
- Říha M, Kubečka J, Vašek M, Seďa J, Mrkvička T, Prchalová M, Matēna J, Hladík M, Čech M, Draštík V, Frouzová J, Hohausová E, Jarolím O, Jůza T, Kratochvíl M, Peterka J, Tušer M (2009) Long-term development of fish populations in the Římov Reservoir. Fisheries Management and Ecology 16: 121–129. https://doi.org/10.1111/j.1365-2400.2008.00650.x
- Rochet MJ, Trenkel VM (2003) Which community indicators can measure the impact of fishing? A review and proposals. Canadian Journal of Fisheries and Aquatic Sciences 60(1): 86–99. https://doi.org/10.1139/f02-164
- Rolfe P (2010) Crock of Gold: Seeking the Crucian Carp. M Press (Media) Ltd, 288 pp.
- Rylková K, Kalous L, Bohlen J, Lamatsch DK, Petrtýl M (2013) Phylogeny and biogeographic history of the cyprinid fish genus Carassius (Teleostei: Cyprinidae) with focus on natural and anthropogenic arrivals in Europe. Aquaculture (Amsterdam, Netherlands) 380–383: 13–20. https://doi.org/10.1016/j.aquaculture.2012.11.027
- Sakai AK, Allendorf FW, Holt JS, Lodge DM, Molofsky J, With KA, Baughman S, Cabin RJ, Cohen JE, Ellstrand NC, McCauley DE, O’Neil P, Parker IM, Thompson JN, Weller SG (2001) The population biology of invasive species. Annual Review of Ecology and Systematics 32(1): 305–332. https://doi.org/10.1146/annurev.ecolsys.32.081501.114037
- Sayer CD, Copp GH, Emson D, Godard MJ, Zięba G, Wesley KJ (2011) Towards the conservation of crucian carp Carassius carassius: Understanding the extent and causes of decline within part of its native English range. Journal of Fish Biology 79(6): 1608–1624. https://doi.org/10.1111/j.1095-8649.2011.03059.x
- Sayer CD, Emson D, Patmore IR, Greaves HM, West WP, Payne J, Davies GD, Tarkan AS, Wiseman G, Cooper B, Grapes T, Cooper G, Copp GH (2020) Recovery of the crucian carp Carassius carassius (L.): Approach and early results of an English conservation project. Aquatic Conservation 30(12): 2240–2253. https://doi.org/10.1002/aqc.3422
- Shin YJ, Rochet MJ, Jennings S, Field JG, Gislason H (2005) Using size-based indicators to evaluate the ecosystem effects of fishing. ICES Journal of Marine Science 62(3): 384–396. https://doi.org/10.1016/j.icesjms.2005.01.004
- Smartt J (2007) A possible genetic basis for species replacement: Preliminary results of interspecific hybridisation between native crucian carp Carassius carassius (L.) and introduced goldfish Carassius auratus (L.). Aquatic Invasions 2(1): 59–62. https://doi.org/10.3391/ai.2007.2.1.7
- Šmejkal M, Kalous L, Velenský P (2021) Zachraň Karase! - Zachrankarase.cz. Save the crucian carp. https://zachrankarase.cz/ [in Czech]
- Šmejkal M, Bartoň D, Duras J, Horký P, Muška M, Kubečka J, Pfauserová N, Tesfaye M, Slavík O (2023) Living on the edge: Reservoirs facilitate enhanced interactions among generalist and rheophilic fish species in tributaries. Frontiers in Environmental Science 11: 1099030. https://doi.org/10.3389/fenvs.2023.1099030
- Szczerbowski JA (2002) Carassius auratus (Linnaeus, 1758). In: Banarescu P, Paepke HJ (Eds) The freshwater fishes of Europe, Vol. 5/III, Cyprinidae 2. AULA-Verlag, Wiesbaden, 5–41.
- Tapkir S, Boukal D, Kalous L, Bartoň D, Souza AT, Kolar V, Soukalová K, Duchet C, Gottwald M, Šmejkal M (2022) Invasive gibel carp (Carassius gibelio) outperforms threatened native crucian carp (Carassius carassius) in growth rate and effectiveness of resource use: Field and experimental evidence. Aquatic Conservation 32(12): 1901–1912. https://doi.org/10.1002/aqc.3894
- Tapkir S, Thomas K, Kalous L, Vašek M, Meador TB, Šmejkal M (2023) Invasive gibel carp use vacant space and occupy lower trophic niche compared to endangered native crucian carp. Biological Invasions 25(9): 2917–2928. https://doi.org/https://doi.org/10.1007/s10530-023-03081-9
- Tarkan AS, Almeida D, Godard MJ, Gaygusuz Ö, Rylands M, Sayer CD, Zieba G, Copp GH (2016) A review and meta-analysis of growth and life-history traits of a declining European freshwater fish, crucian carp Carassius carassius. Aquatic Conservation 26(1): 212–224. https://doi.org/10.1002/aqc.2580
- Tarkan AS, Mol O, Aksu S, Köse E, Kurtul I, Başkurt S, Haubrock PJ, Balzani P, Çınar E, Britton JR, Oztopcu-Vatan P, Emiroğlu Ö (2023) Phenotypic responses to piscivory in invasive gibel carp populations. Aquatic Sciences 85(3): 75. https://doi.org/10.1007/s00027-023-00974-8
- Thomas K, Brabec M, Tapkir S, Gottwald M, Bartoň D, Šmejkal M (2023) Sampling bias of invasive gibel carp and threatened crucian carp: Implications for conservation. Global Ecology and Conservation 48: e02718. https://doi.org/10.1016/j.gecco.2023.e02718
- Thornton TF, Scheer AM (2012) Collaborative Engagement of Local and Traditional Knowledge and Science in Marine Environments: A Review. Ecology and Society 17(3): art8. https://doi.org/10.5751/ES-04714-170308
- Tóth J (1976) A brief account on the presence of the silver crucian (Carassius auratus gibelio Bloch 1783) in the Hungarian section of the Danube. Section Biologica, 219–220.
- van den Heuvel L, Rönnbäck P (2023) What you see isn’t always what you get: On how anglers’ fish stock perceptions are influenced by motivations, satisfaction and engagement. Fisheries Research 258: 106519. https://doi.org/10.1016/j.fishres.2022.106519
- van der Veer G, Nentwig W (2015) Environmental and economic impact assessment of alien and invasive fish species in Europe using the generic impact scoring system. Ecology Freshwater Fish 24(4): 646–656. https://doi.org/10.1111/eff.12181
- Vejřík L, Vejříková I, Kočvara L, Blabolil P, Peterka J, Sajdlová Z, Jůza T, Šmejkal M, Kolařík T, Bartoň D, Kubečka J, Čech M (2019) The pros and cons of the invasive freshwater apex predator, European catfish Silurus glanis, and powerful angling technique for its population control. Journal of Environmental Management 241: 374–382. https://doi.org/10.1016/j.jenvman.2019.04.005
- Venturelli PA, Hyder K, Skov C (2017) Angler apps as a source of recreational fisheries data: opportunities, challenges and proposed standards. Fish and Fisheries 18(3): 578–595. https://doi.org/10.1111/faf.12189
- Vinterstare J, Bronmark C, Nilsson PA, Langerhans RB, Chauhan P, Hansson B, Hulthen K (2023) Sex matters: Predator presence induces sexual dimorphism in a monomorphic prey, from stress genes to morphological defences. Evolution 77(1): 304–317. https://doi.org/10.1093/evolut/qpac030
- Wang L, Akritas MG, Van Keilegom I (2008) An ANOVA-type nonparametric diagnostic test for heteroscedastic regression models. Journal of Nonparametric Statistics 20(5): 365–382. https://doi.org/10.1080/10485250802066112
- Wickham H (2016) ggplot2: Elegant Graphics for Data Analysis. Springer-Verlag New York. https://doi.org/10.1007/978-3-319-24277-4
- Wilde GR, Pope KL (2004) Anglers’ probabilities of catching record-size fish. North American Journal of Fisheries Management 24(3): 1046–1049. https://doi.org/10.1577/M03-100.1
- Wood SN (2001) mgcv: GAMs and generalized ridge regression for R. R News.
- Wood SN (2017) Chapman and Hall/CRC Generalized Additive Models. Chapman and Hall/CRC. https://doi.org/10.1201/9781315370279
- World Bank (2002) Hidden harvest: the global contribution of capture fisheries (English). World Bank Group, Washington, D.C.
- Wouters J, Janson S, Lusková V, Olsén KH (2012) Molecular identification of hybrids of the invasive gibel carp Carassius auratus gibelio and crucian carp Carassius carassius in Swedish waters. Journal of Fish Biology 80(7): 2595–2604. https://doi.org/10.1111/j.1095-8649.2012.03312.x
Supplementary material
Individual data points
Data type: docx