





(C) 2012 Clare Aslan. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Seed disperser preferences may mediate the impact of invasive, non-native plant species on their new ecological communities. Significant seed disperser preference for invasives over native species could facilitate the spread of the invasives while impeding native plant dispersal. Such competition for dispersers could negatively impact the fitness of some native plants. Here, we review published literature to identify circumstances under which preference for non-native fruits occurs. The importance of fruit attraction is underscored by several studies demonstrating that invasive, fleshy-fruited plant species are particularly attractive to regional frugivores. A small set of studies directly compare frugivore preference for native vs. invasive species, and we find that different designs and goals within such studies frequently yield contrasting results. When similar native and non-native plant species have been compared, frugivores have tended to show preference for the non-natives. This preference appears to stem from enhanced feeding efficiency or accessibility associated with the non-native fruits. On the other hand, studies examining preference within existing suites of co-occurring species, with no attempt to maximize fruit similarity, show mixed results, with frugivores in most cases acting opportunistically or preferring native species. A simple, exploratory meta-analysis finds significant preference for native species when these studies are examined as a group. We illustrate the contrasting findings typical of these two approaches with results from two small-scale aviary experiments we conducted to determine preference by frugivorous bird species in northern California. In these case studies, native birds preferred the native fruit species as long as it was dissimilar from non-native fruits, while non-native European starlings preferred non-native fruit. However, native birds showed slight, non-significant preference for non-native fruit species when such fruits were selected for their physical resemblance to the native fruit species. Based on our review and case studies, we propose that fruit characteristics of native plant communities could dictate how well a non-native, fleshy-fruited plant species competes for dispersers with natives. Native bird preferences may be largely influenced by regional native fruits, such that birds are attracted to the colors, morphology, and infructescence structures characteristic of preferred native fruits. Non-native fruits exhibiting similar traits are likely to encounter bird communities predisposed to consume them. If those non-natives offer greater fruit abundance, energy content, or accessibility, they may outcompete native plants for dispersers.
Catharus guttatus, feeding preference, frugivory, Sturnus vulgaris, Turdus migratorius
Invasive non-native plant species may exert a range of impacts on native communities. They can alter resource availability (e.g.,
Competition with invasive plants may further impact native communities by altering interactions between native species. Invaders can transform food web structure, for example, by outcompeting high-quality native food plants and thereby diminishing an herbivore’s resources (
Among introduced species, fleshy-fruited plants adapted to animal-mediated seed dispersal are often considered high-risk for invasiveness (
Animal-dispersed invasive plants can have the same community impacts as other invasives, forming monospecific thickets (
There have been a few attempts in localized systems to determine whether competition for dispersers occurs. Gosper et al. (2006) examined vertebrate-mediated dispersal of native plant species growing with and without invasive Chrysanthemoides monilifera, but found no effect of the invasive on native dispersal.
Since preference may play a key role in such competition, we combined a literature review, exploratory meta-analysis, and two case study aviary experiments to address the question: Under what circumstances do seed dispersers demonstrate preference for non-native fruits? We began by examining a number of studies that have probed the causes of fleshy-fruited invasion and identified characteristics that appear to promote animal-mediated dispersal of the invasive species in question. These studies do not directly compare native and non-native seed dispersal, but shed light on factors that may be pivotal in seed disperser preference. Additionally, a limited number of studies have directly examined frugivore preference for native vs non-native fruits (Table 1). These studies have fallen into two categories: a small set of experiments comparing disperser preference for native and non-native species selected for their similarity; and somewhat more numerous studies comparing disperser preference for native and non-native species in an existing suite of co-occurring species, with no attempt to control species similarity. We conducted a small-scale meta-analysis on comparative studies containing sufficient quantitative information to examine relative disperser preferences for non-native vs. native species. Implications of resulting preference data varied qualitatively depending on study category. We illustrate the contrasting outcomes of the two approaches by including here the results of two aviary case studies we conducted in which we compared bird preferences for native and non-native species when fruits were controlled for similarity and when they were not. In combination, our review and case studies suggest that the regional native fruit assemblage is likely to strongly influence frugivore preferences, and that the degree to which an introduced species monopolizes dispersal services may depend on its superficial similarity to native species.
Studies that have directly compared seed disperser preferences for native vs. non-native fruits. Effect size, used in meta-analysis calculation, is the natural log of the response ratio, calculated as the proportion of non-native fruit consumed to the proportion of native fruit consumed. Studies with insufficient quantitative data were not included in the meta-analysis, although their qualitative results are discussed in the review. Superior feeding efficiency reflects author interpretations within each study and may include factors such as larger fruit size, higher fruit energy or nutritional content, higher fruit flesh to seed ratio, larger infructescences, larger plants, and more accessible fruits. † Approach distinguishes between studies that deliberately matched surface fruit characteristics such as color and fruit type (= “matched”) and studies that examined existing suites of fruits without controlling for similarity (= “unmatched”).
| Study | Disperser | Effect size (lnRR) | Offers superior feeding efficiency | |
|---|---|---|---|---|
| Approach †: Unmatched | ||||
|
|
Birds | Insufficient quantitative data | Non-native | |
|
|
Birds | -0.06 | Native | |
|
| Ferreras et al. 2008 | Birds | Insufficient quantitative data | Native |
|
|
Birds | 0.04 | No clear superior | |
|
|
Birds | 0.14 | No clear superior | |
|
|
Birds | -0.12 | Not discussed | |
|
|
Birds | -0.20 | Not discussed | |
|
|
Ants | -0.33 | Non-native | |
|
|
Birds | -0.82 | No clear superior | |
|
|
Birds | Insufficient quantitative data | Native | |
|
|
Birds | Insufficient quantitative data | Native | |
| Our experiment #1, this paper | Birds | -0.62 | No clear superior | |
| Approach †: Matched | ||||
|
| Greenberg et al. 2001 | Birds | -0.14 | Not discussed |
|
|
Birds | -1.84 | Native | |
|
| LaFleur et al. 2007 | Birds | 0.95 | Non-native |
|
|
Birds | Insufficient quantitative data | Non-native | |
| Our experiment #2, this paper | Birds | 0.42 | Not explored | |
Many studies in invasion biology are intended to elucidate the causes of ongoing invasions by particular species. Among fleshy-fruited invaders, fruit attractiveness to frugivore dispersers is frequently examined. Characteristics such as color, fruit size, infructescence structure, nutritional content, and pulp to seed ratio are considered general determinants of fruit attractiveness to frugivores (
The small number of experiments that have examined frugivore preferences for native vs. non-native fruits that are similar in surface characteristics such as color and structure (e.g., both with red berries) have more often found preference for non-native than for native fruiting species (Table 1). Authors have suggested that frugivores use fine- scale differences to distinguish between similar food items. In two out of three aviary choice tests among color-matched pairs, European starlings and American robins preferred non-native over native fruits (
When combinations of native and invasive species in existing plant communities have been examined for frugivore preference, most studies have detected lack of preference or preference for native fruits (Table 1). No significant preference for either native or non- native fruits was observed when fruit removal by birds of two non-native and five native tree species co-occurring in Argentina was examined (
Because such a small number of studies have directly compared frugivore preferences for native vs. non-native fruits, a quantitative examination of combined study results has limited power. Therefore, although we conducted an exploratory meta-analysis to evaluate overall trends, we consider meta-analysis results preliminary and urge further study before general conclusions regarding frugivore preferences may be drawn.
Meta-analysis methodsFor our meta-analysis, we used the response ratio to compare effect sizes between studies (
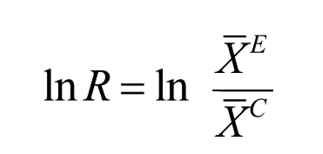
where X¯E is the weighted mean of the proportion of non-native fruits consumed and X¯C is the weighted mean of the proportion of native fruits consumed. Using this metric, a negative mean effect size with a 95% bias-corrected confidence interval excluding zero indicates that native fruits are consumed significantly more than non-native fruits. A positive mean effect size with a 95% bias-corrected confidence interval excluding zero indicates that non-native fruits are consumed significantly more than native fruits. We used a random-effects model and generated confidence intervals using 5000 data randomizations (
The overall meta-analysis detected no significant difference in frugivore consumption of native vs. non-native fruits (mean effect size = -0.21, 95% bias-corrected confidence interval -0.35 to 0.07, n = 12 studies). However, categorical analysis detected differences between studies examining matched vs. unmatched fruits. For studies in which fruits were matched by external characteristics, there was no significant difference in frugivore preference between native and non-native fruits, but a trend toward preference for non-native fruits was visible (mean effect size = 0.37, 95% bias-corrected confidence interval -0.16 to 0.91, n = 4 studies). For studies in which no attempt was made to match fruits, frugivores demonstrated significant preference for native fruits (mean effect size = -0.31, 95% bias-corrected confidence interval -0.37 to -0.03, n = 8 studies). Again, the number of studies available for each category was small enough that these meta-analysis calculations had low power and results should be treated with caution.
Case study: Bird preferences among fleshy-fruited species in northern CaliforniaWe conducted two aviary feeding preference experiments using wild-caught birds and fleshy-fruited plants in northern California. Our results illustrate the importance of fruit similarity in experimental conclusions.
Field-based foraging observations on fleshy fruits in California demonstrated higher visitation and fruit consumption rates for native Heteromeles arbutifolia than for co-occurring non-natives Olea europaea, Ligustrum lucidum, and Triadica sebifera (
The four non-native plants used in the first experiment are widely planted in the Sacramento Valley of California, co-occurring with the native species in many locations. Olea europaea L. (European olive, Oleaceae) produces racemes of large (ranging from 6–21 mm width), dark-purple drupes with high oil content and has become invasive in Australia (
The bird species used in this experiment were selected to represent different functional guilds identified in field observations (
Ten robins were captured with mist nets in Butte County, CA, in December, 2007. Nineteen starlings were captured with walk-in traps in Marin County, CA, in November, 2008. Seven hermit thrushes were captured with mist nets in Butte and Yolo Counties, CA, in January, 2009. All birds were returned to open-air cages at the University of California, Davis. Cages measured 2.4 m tall, 1.5 m wide, and 3.8 m long. A single bird was housed in each cage and opaque green shade cloth was attached to walls between cages to prevent birds from observing and mimicking one another. Cages were roofed with metal sheeting to shelter birds from precipitation but were open at either end to allow natural sunlight and airflow/temperature. Birds were kept in cages for 10 days following capture to enable them to adapt to cage conditions. During this period, birds were provided ad libitum a maintenance diet with a banana/soy protein base (
For choice tests, experimental fruits were hung from weighted fishing line attached to cage ceilings. Fresh infructescences were collected from plants during the morning of each trial and were suspended from fishing line by looping the line around the infructescence stems. A separate line was used for each plant species so that the infructescences were presented to birds in a row across one end of the cage, separated from one another by approximately 30 cm. A long perch was hung in front of the row of fruits, allowing the bird easy access to all fruits. The fishing line allowed infructescences to bounce naturally as the bird foraged, mimicking branch movement in the wild. Whole infructescences were used so that the appearance, relative abundance, and arrangement of fruits reflected those encountered in the field. There was no attempt to artificially enhance similarity between plant species by controlling for fruit size or per-infructescence fruit abundance. Size differences between fruits are so great that any attempt to equalize mass, for example, would require a given trial to utilize a very small number of Olea europaea fruits relative to the number of Ligustrum lucidum fruits, skewing food item abundance dramatically. Whole infructescence use resulted in Ligustrum lucidum being the most numerous fruit offered, while Olea europaea was usually offered in the greatest mass. Statistical analyses took into account initial availability of each fruit species per trial. At the end of each trial, fruits of all four species remained in all bird cages, demonstrating that quantities initially provided were sufficient to allow birds to make choices throughout the duration of each trial.
Choice trials were conducted as follows: Each phase of choice trials lasted six days. Just before daybreak on each day, one ripe (containing only fruits visibly as bright in color as any available on regional source trees) infructescence of each of the four study species was placed in each cage. Birds had fasted overnight. Infructescences were hung in random order from the fishing lines so that the arrangement differed from day to day. All birds were offered all fruits each day. All fruits were counted prior to their placement in the cages. Birds were allowed to forage freely for two hours. Birds showed evidence of hunger during this time, since most birds foraged, but not starvation (on some days, a minority of birds did not forage during the two-hour period). At the end of this period, all infructescences were removed and maintenance food replaced in cages. The final numbers of fruits per infructescence were counted to determine how many fruits had been removed. Cage floors were inspected and any whole fruits that had been dropped or had fallen from infructescences were collected so that actual numbers of fruits consumed could be calculated. Samples of fruits of each species were then weighed so that approximate masses of fruits available and consumed could be estimated. The same fruit species were offered on each of the six days, and these days were considered subsamples for statistical calculations. This minimized the effect of weather and other factors that may have caused variation in birds’ daily feeding behavior.
During the second experimental phase, the fruit species most preferred by each bird species was withheld in order to determine preference hierarchy among remaining fruit species (to force some birds to choose between species they had previously ignored). Again, results from each day within this phase were considered subsamples for calculation of within-phase preference. Because robins and hermit thrushes preferred the native Heteromeles arbutifolia during phase 1, a third experimental phase included the native fruit once again in the experimental array so that birds had all four options. This was to ensure that preferences for the native in phase 1 were not due simply to familiarity with the native fruit, since it is more widespread in northern California than are the target non-natives, which are still clustered around urban and agricultural areas. Because birds had consumed non-native fruits during phase 2, when no natives were available, we hypothesized that neophobia or any similarly transient cause of preference would be purged by phase 2. Phase 3 thus elucidated the robustness of the initial preference for the native fruiting species, determining whether birds maintained that preference even after demonstrated familiarity with non-native fruits.
Preference hierarchy among plants selected for resemblanceIn the second experiment, seven hermit thrushes (captured and maintained following the same methods and facilities described above) were offered fruits of four species selected for their resemblance to one another in the surface characteristics of fruit color, shape, and size: native Heteromeles arbutifolia and non-natives Cotoneaster pannosus, Photinia x fraseri, and Pyracantha angustifolia. All three of these non-native species are in the family Rosaceae and produce panicles of small red pomes. Cotoneaster pannosus is considered a potentially invasive species in California due to its invasiveness elsewhere and observed spreading in natural areas (
Number of fruits consumed is more relevant to seed dispersal than is mass consumed since the number of fruits translates directly to the number of seeds that could be dispersed. For simplicity, therefore, figures and tables include only results by number. All statistical tests were performed for mass as well, but results did not differ qualitatively from results by number.
Treating the days within each experimental phase as subsamples, we calculated mean numbers available and consumed of each fruit species by each bird. We used these averages to calculate the selection index (wi) of each fruit species by individual bird: wi = oi/pi, where oi = the proportion of species i in the diet, and pi = the proportion of species i available in the environment (
For the first experiment, analysis of variance revealed significant feeding preferences by selection index for all bird species in all experimental phases (Table 2, Figure 1). Robins preferred native Heteromeles arbutifolia to other test fruits when the native was available. When Heteromeles arbutifolia was withheld, they preferred Olea europaea (Figure 1a). On average, starlings preferred Olea europaea to other experimental fruit species. When Olea europaea was withheld, starlings avoided Triadica sebifera but preferred Ligustrum lucidum and Heteromeles arbutifolia equally (Figure 1b). Hermit thrushes preferred Heteromeles arbutifolia to other fruits (both first and third experimental phases). When the native was withheld, they preferred Olea europaea to other fruits with Ligustrum lucidum as a close second (Figure 1c). Triadica sebifera was consumed least by all birds during all phases (Figure 1).
Results of analyses of variance comparing fruit species selection indices (wi) separated by bird species and by experimental phase. Individual birds were treated as blocks for analysis. Data were square- root transformed to meet assumptions of homogeneity of variance.
| Bird species |
Experimental phase |
Source | DF | F ratio | Prob > F |
|---|---|---|---|---|---|
| American robin (Turdus migratorius) | 1 | Fruit | 3 | 26.82 | <0.0001 |
| Bird (Block) | 9 | 0.4035 | 0.9222 | ||
| 2 | Fruit | 2 | 13.68 | 0.0002 | |
| Bird (Block) | 9 | 0.6943 | 0.7058 | ||
| 3 | Fruit | 3 | 55.07 | <0.0001 | |
| Bird (Block) | 9 | 0.6974 | 0.7055 | ||
| European starling (Sturnus vulgaris) | 1 | Fruit | 3 | 13.68 | <0.0001 |
| Bird (Block) | 18 | 0.4404 | 0.9711 | ||
| 2 | Fruit | 2 | 7.95 | 0.0014 | |
| Bird (Block) | 18 | 0.6810 | 0.8064 | ||
| Hermit thrush (Catharus guttatus) | 1 | Fruit | 3 | 33.82 | <0.0001 |
| Bird (Block) | 6 | 0.6470 | 0.6921 | ||
| 2 | Fruit | 2 | 4.35 | 0.0380 | |
| Bird (Block) | 6 | 0.4374 | 0.8403 | ||
| 3 | Fruit | 3 | 9.97 | 0.0004 | |
| Bird (Block) | 6 | 0.0911 | 0.9965 |
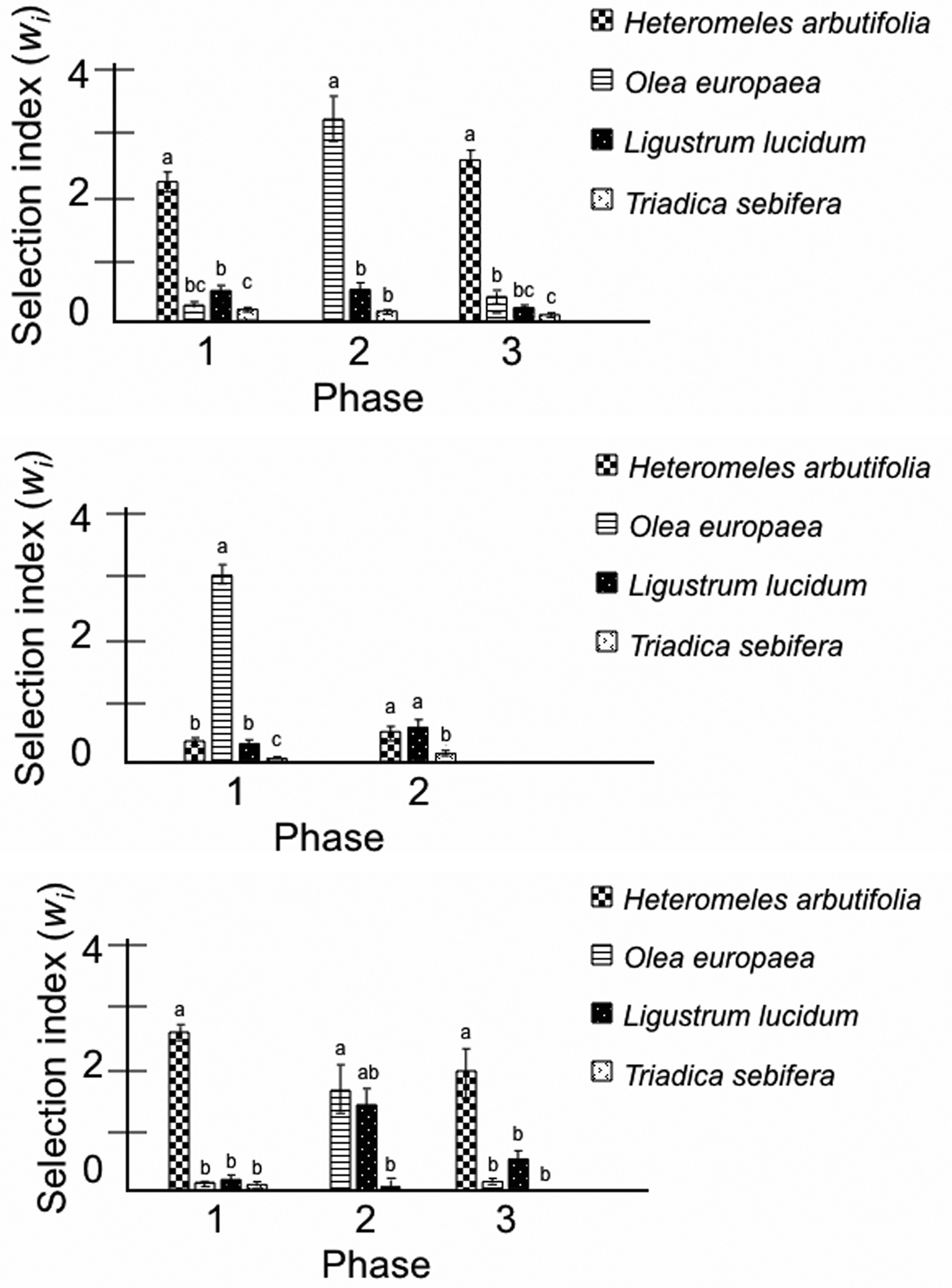
Mean (± SE) selection indices (wi) by bird species and by experimental phase. Selection index values >1.0 indicate a preferred food item (consumed at a rate exceeding its availability). Analyses of variance with individual birds designated as blocks compared preferences within each experimental phase. Means within each ANOVA were compared using Tukey HSD means separation. Within each experimental phase, bars not designated by the same letter are significantly different at a = 0.05. a American robins (Turdus migratorius) b European starlings (Sturnus vulgaris) c Hermit thrushes (Catharus guttatus)
During the second experiment, when experimental fruits were selected for their similarity, hermit thrush selection indices displayed little preference structure (Table 3). Analysis of variance results showed no overall significant difference in feeding preference among the four red-fruited species (Figure 2; a = 0.05), although raw numbers demonstrated greater consumption of all three non-native fruits than of the native Heteromeles arbutifolia.
Selection indices (wi) for similarity experiment using red fruit and testing preferences of hermit thrushes (Catharus guttatus). Selection index values greater than 1.0 indicate preference by hermit thrush individuals for a food item, and these values are underlined. Values are presented for number of fruits as selection indices ± SE.
| Bird | Pyracantha angustifolia | Cotoneaster pannosus | Photinia x fraseri | Heteromeles arbutifolia |
|---|---|---|---|---|
| A | 0.12 ± 0.21 |
0.36 ± 0.31 |
2.51 ± 0.37 |
0.50 ± 0.32 |
| B | 0.26 ± 0.32 |
1.30 ± 0.57 |
1.32 ± 0.44 |
0.87 ± 0.48 |
| C |
1.94 ± 0.70 |
0.98 ± 0.54 |
0.47 ± 0.36 |
0.82 ± 0.49 |
| D |
1.12 ± 1.23 |
0.97 ± 1.06 |
0.80 ± 0.88 |
1.21 ± 1.33 |
| E |
2.95 ± 0.73 |
0.21 ± 0.26 |
0.96 ± 0.41 |
0.26 ± 0.29 |
| F | 0.50 ± 0.36 |
0.40 ± 0.28 |
2.33 ± 0.32 |
0.36 ± 0.23 |
| G | 0.40 ± 0.39 |
0.93 ± 0.52 |
1.60 ± 0.40 |
0.66 ± 0.43 |
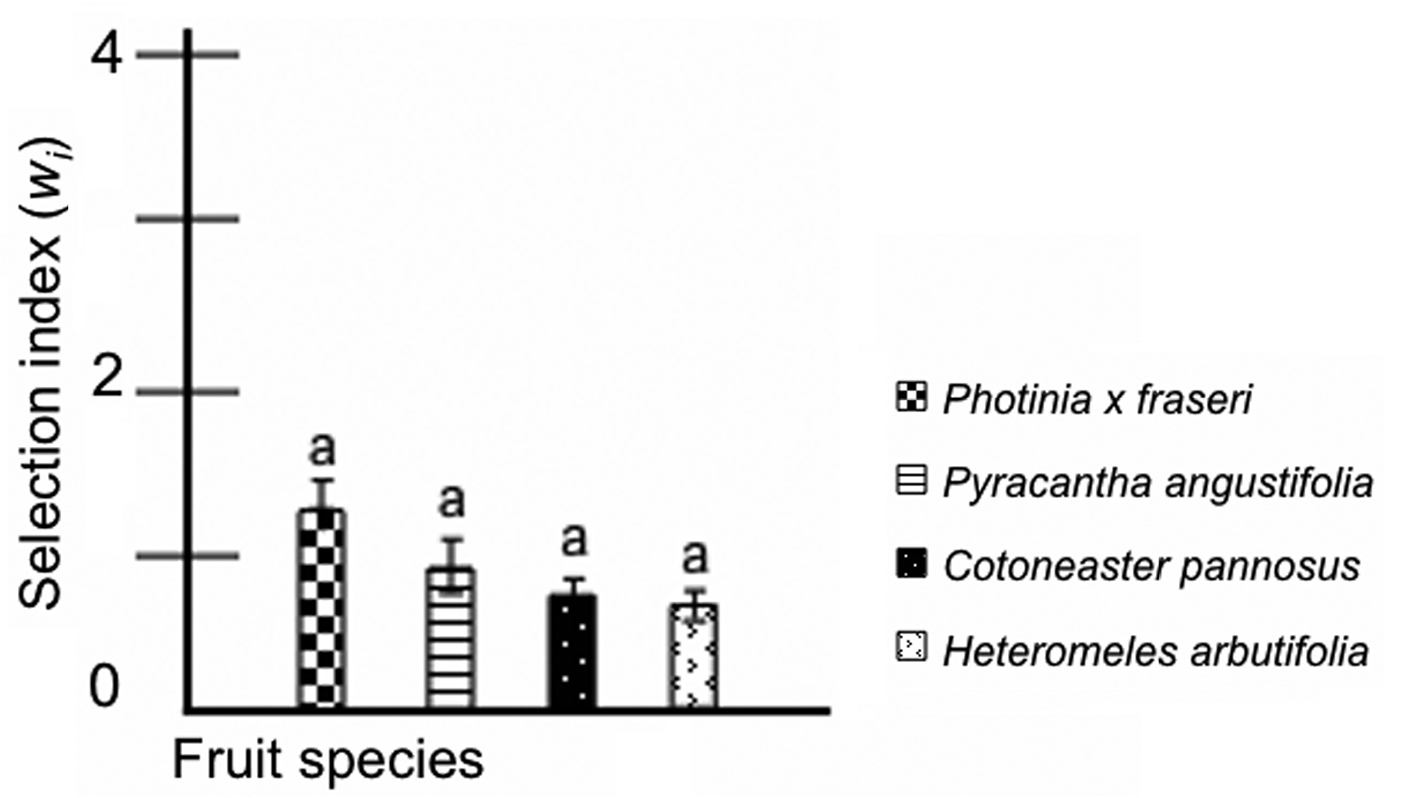
Hermit thrush (Catharus guttatus) fruit similarity experiment selection indices by fruit species. Mean selection indices were compared using analysis of variance, with individual bird as block. Means within each ANOVA were compared using Tukey HSD means separation. Within each experimental phase, bars not designated by the same letter are significantly different at a = 0.05.
If native frugivores prefer introduced fruiting species in a given region, seed dispersal services are likely to be monopolized to some degree by the non-native plant species and withheld from native plants. Although the number of studies that have explored such dispersal competition is quite low, our review and meta-analysis of their results and our case study experiments indicate that native fruit characteristics may play an important role in determining frugivore preference. When frugivore preference is evaluated in the context of the full existing suite of fleshy-fruited plants, with no attempt to artificially enhance similarity between species, frugivores tend to prefer native fruits. However, when non-natives that are similar to natives in surface appearance have been examined, frugivores have shown an inclination, often slight, to prefer the non-native fruits. In published literature, preference for non-natives appeared to result from larger crop sizes, higher per-fruit energy content, or larger fruits relative to seed mass, implying that such species are preferred because the reward is likely greater for a given frugivore visit. Thus, frugivores in these studies seem to demonstrate general attraction to certain characteristics typical of native fruits (such as color or shape), and it is within sets of species sharing these characteristics that more subtle preferences favoring non-native species appear.
In our case study experiments, for example, both native bird species preferred the native red berry even above Ligustrum lucidum with its very large crop sizes of dark purple berries, although the two are extremely similar nutritionally. When the native fruit was contrasted against non-native, red-berried species offered to hermit thrushes, however, this strong preference for the native fruit disappeared. Birds that had displayed highly significant preference for native fruits displayed no significant preference structure when similar fruits were introduced. The non-native bird species, on the other hand, showed no preference at any time for the native species. The native bird fauna appears to maintain a robust preference for red-berried species, which could be the result of loose coevolution between the birds and fruits in the area: since the most available native fruit during the winter season is red, a preference for that color among native birds is unsurprising. When non-native species producing red berries are introduced to this system, they are likely preadapted to ready winter seed dispersal. As long as they produce larger or more accessible crops, they may draw dispersers away from the native species. By contrast, fruits of different color or structure from the native appear less preferred, likely because they lack such preadaptation. Under this scenario, native fruit characteristics appear to play a key role in determining whether native and non-native fruits will compete for dispersers. Introduced species sharing these characteristics (red berries) are most likely to exert a competitive impact. An important consideration is that bird adaptation to prefer a given native fruit may arise because that native fruit is a particularly suitable food source, for example contributing to higher bird fitness by offering high nutritional value. Decline in such fruit species through a competitive effect may therefore impact native bird populations.
Non-native frugivores may also affect seed dispersal. Such species have enhanced the dispersal of native species in some systems (
The review, meta-analysis, and aviary experiments examined here suggest that frugivores are most likely to favor non-native fruits if they resemble preferred native fruits, at least on the surface. In several instances, frugivores have shown preference for non-native species that are selected to resemble native species in fruit color or type. When such introduced species offer a larger number of fruits per plant or relatively higher quantity of digestible material per fruit, they appear to attract more frugivory than the natives and thus have the potential to display a competitive edge over native fleshy fruits. When non-native species differ from natives in superficial characteristics such as fruit color or type, studies show a tendency for frugivores to prefer native species or at least to feed opportunistically. This pattern implies that non-native fruits are most likely to encounter preferential frugivory and thus to compete for dispersers with native fruits if they resemble those native fruits and offer enhanced feeding efficiency. When these conditions are met, competition for dispersers and eventual decline in the dispersal of native species may occur following fleshy-fruiting plant introductions.
If it is indeed widespread across geographies, such competition has implications both before and after plant introductions. When new plants are considered for a region, as horticultural, agricultural, or landscaping introductions, they may be most likely to encounter frequent dispersal and to exert competitive impact if they resemble preferred native plants and overlap with natives in fruiting phenology. Along with other known invasive impacts of such species, dispersal competition should be considered a potential additional impact of their introduction, and should be taken into account. Control of such species in favor of the native should be considered.
To date, the number of studies directly addressing seed disperser preference for native vs. non-native fruits is very low, and study designs differ considerably. A greater number of such studies will be necessary in order to evaluate with confidence how widespread these patterns may be. In addition, we recommend use of standardized methodologies (such as those described in
Comments from T. Hahn, M. Holyoak, D. Levey, and A. Loayza greatly improved this manuscript. We are deeply grateful to J. Ellsworth for aviary construction and design and experimental setup ideas. We thank T. Hahn for equipment loans and bird maintenance suggestions and M. Holyoak for experimental design advice. The Department of Wildlife, Fish, and Conservation Biology at the University of California, Davis, provided aviary facilities; thanks to W. P. Gorenzel for facility assistance. W. Ferrier provided veterinary care and advice. We thank the Big Chico Creek Ecological Reserve, the Yolo Audubon Society, and the County of Solano for permission to mist net on their lands. Thanks to B. Hull, who captured the starlings used in this study. Mist net training and assistance were provided by J. Cornelius, R. Kelsey, K. Richman, and D. Garcia. Thanks to E. Alonzo, E. Armstrong, A. Aslan, R. Boothby, S. Bouret, M. Bower, L. Brizzolara, S. Chin-Shar Chu, M. Chinoraks, R. DeRuvo, P. Ellsworth, H. Jones, S. Krause, A. Lewis, C. Liang, R. McKee, K. Moore, J. Piekut, and A. Rauth, who assisted with bird care and counted thousands of fruits. This work was supported by a Montana State Center for Invasive Plant Management Seed Money Grant award and a research grant from the UC Davis Biological Invasions IGERT (NSFDGE#0114432). C.E.A. was supported by a National Science Foundation Graduate Research Fellowship. All work was performed in accordance with UCD Institutional Animal Care and Use Committee permit number 12559.
Nutrient content of the four focal fruit species. Values are for fruit pulp, the fruit portion assimilated in most cases by frugivorous birds, which usually defecate or regurgitate seeds. Nutrient content analyses were conducted by a commercial food analysis company (National Food Laboratories, Inc., Livermore, CA).
| Nutritional component |
Heteromeles arbutifolia | Olea europaea | Ligustrum lucidum | Triadica sebifera |
|---|---|---|---|---|
| Fructose (%) | 0.62 | 0.11 | 2.77 | <0.25 |
| Glucose (%) | 5.06 | 1.06 | 5.80 | <0.25 |
| Lactose (%) | <0.25 | <0.25 | <0.25 | <0.25 |
| Maltose (%) | <0.25 | <0.25 | <0.25 | <0.25 |
| Sucrose (%) | <0.25 | <0.25 | 0.63 | 2.27 |
| Total Sugars (%) | 5.88 | 1.67 | 9.71 | 2.27 |
| Ash (%) | 1.14 | 2.28 | 1.84 | 2.33 |
| Calories (cal/100g) | 134.07 | 346.21 | 95.07 | 1038.46 |
| Carbohydrates (%) | 33.28 | 22.04 | 25.75 | 3.16 |
| Moisture (%) | 64.91 | 45.78 | 72.24 | 28.01 |
| Protein (%) | 0.67 | 2.18 | 0.17 | <0.25 |
| Total Fat (%) | <0.25 | 27.71 | <0.25 | 66.5 |