





(C) 2012 Matteo Dainese. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
One of the most robust emerging generalisations in invasion biology is that the probability of invasion increases with the time since introduction (residence time). We analysed the spatial distribution of alien vascular plant species in a region of north-eastern Italy to understand the influence of residence time on patterns of alien species richness. Neophytes were grouped according to three periods of arrival in the study region (1500–1800, 1800–1900, and > 1900). We applied multiple regression (spatial and non-spatial) with hierarchical partitioning to determine the influence of climate and human pressure on species richness within the groups. We also applied variation partitioning to evaluate the relative importance of environmental and spatial processes. Temperature mainly influenced groups with species having a longer residence time, while human pressure influenced the more recently introduced species, although its influence remained significant in all groups.Partial regression analyses showed that most of the variation explained by the models is attributable to spatially structured environmental variation, while environment and space had small independent effects. However, effects independent of environment decreased, and spatially independent effects increased, from older to the more recent neophytes. Our data illustrate that the distribution of alien species richness for species that arrived recently is related to propagule pressure, availability of novel niches created by human activity, and neutral-based (dispersal limitation) processes, while climate filtering plays a key role in the distribution of species that arrived earlier. This study highlights the importance of residence time, spatial structure, and environmental conditions in the patterns of alien species richness and for a better understanding of its geographical variation.
Climate, dispersal limitation, energy, environmental filtering, human pressure, land-use, niche-based processes, propagule pressure
Understanding the factors that determine the spatial distribution of exotic species is a primary objective of invasion ecology (
Stochastic factors, including initial inoculum size, residence time, propagule pressure, and chance events (
Recent advances in biogeographical research indicate that the likelihood of biological invasions at the macro scale might be reasonably well predicted simply from knowledge of climatic condition and human-impact (
Several studies have also suggested that ecosystems or habitats differ considerably in the number of alien species they harbour (
Nonetheless, the patterns of distribution of species are determined by a combination of environmental and spatial processes. Therefore, to understand the determinants of variation in species richness it is important to disentangle the effects of environmental and spatial variables. Species distribution patterns are spatially structured for several reasons: first of all, ecological processes are inherently spatial as they operate between neighbouring individuals; secondly species respond to variations in environmental factors, which are themselves spatially structured, thus inducing spatial dependencies in the distributions of species (
In the present study, we analysed the spatial distribution of alien species in a region of north-eastern Italy characterised by high climatic and land-use heterogeneity to understand the influence of minimum residence time (MRT) on patterns of alien species richness. We conducted analyses with all alien plants occurring in the study region and within separate groups of alien species defined by residence time to evaluate the relative importance of climate, human pressure and landscape within the groups. More specifically, the primary objective was to quantify the relative role of environmental conditions and spatial patterns that could arise from niche-based processes such as environmental filtering (
The study area was the Friuli Venezia Giulia region (north-eastern Italy), an area of 7845 km2 (WGS84: N45°34.5'–46°38.3', E12°18.1'–13°55.1') on the southern border of the European Alps. About 43% of the territory is occupied by mountains, 38% by plains, and the remaining 19% by hills. The Adriatic coast extends for c.150 km, from the mouth of the Tagliamento River in the west to the Slovenian border in the east. The elevation ranges from sea level to 2780 m a.s.l. The local climates vary from sub-Mediterranean conditions in the southeast to alpine conditions in the inner valley. Differences in precipitation were mainly related to orographic effects. Northward, the highest precipitation occurs on the external alpine ridge, where the humid sea air is forced to rise over the mountain range. The average annual rainfall varies from c.1000 mm year-1 along the Adriatic coast to c.3000 mm year-1 in the Julian Prealps. The annual mean temperature is 8.7°C and varies from 1.5°C in theAlps to 14.1°C along the Adriatic coast.
Data on alien plant speciesInformation on the distribution of vascular plants was extracted from the floristic atlas of the Friuli Venezia Giulia region (
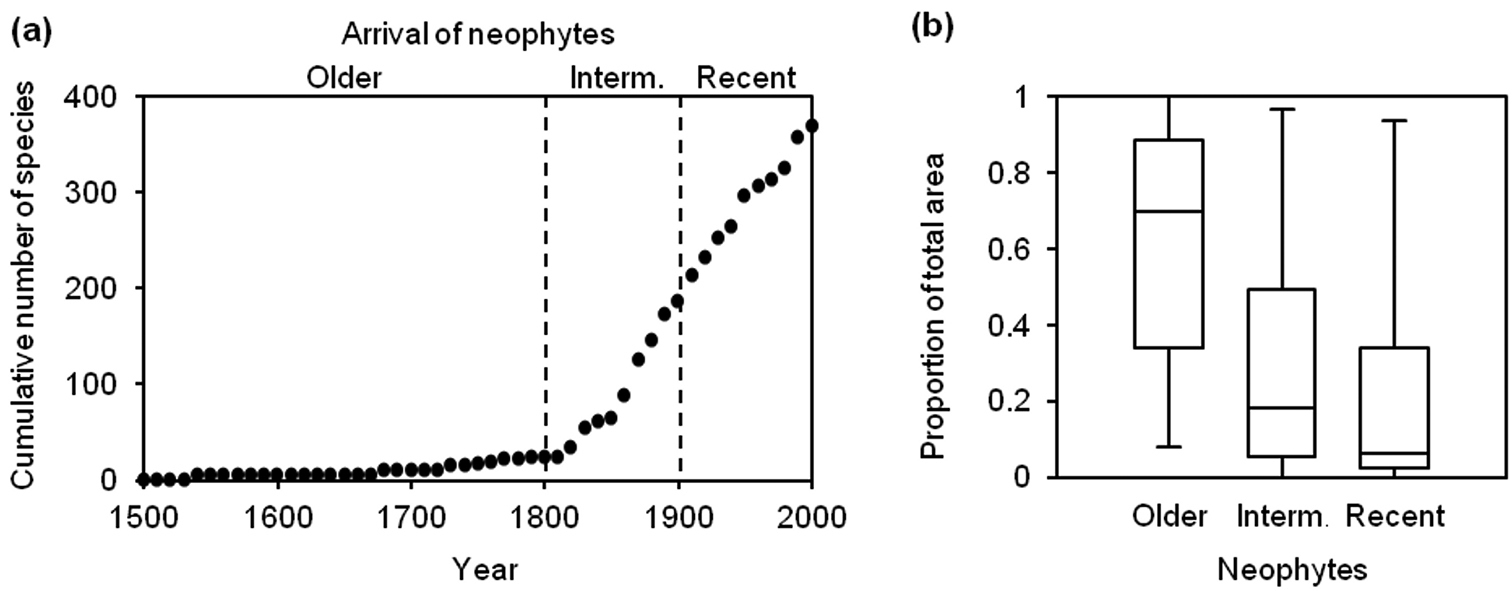
(a) Pattern of accumulation of neophyte species in the Friuli Venezia-Giulia region after the 15th century. Dashed lines divide the curve into three time periods used to differentiate three species groups: 1500–1800, a long period of gradual colonisation (older neophytes); 1800–1900, a first acceleration of the arrivals (intermediate neophytes); > 1900, a second acceleration (recent neophytes). (b) Box and whisker plots of the range sizes as a proportion of the total mapping units of three groups of neophytes differing in minimum residence time.
The dates of the first records in the study region were assembled from the checklist of Italian alien species (
We used the same grid to generate a series of explanatory variables quantifying climate, human-impact, and composition of the landscape. For each group we selected a first set of 35 variables of interest (Appendix S1).
For climatic variables, we considered annual precipitation (PREC) as an indicator of water availability and annual mean temperature (TEMP) as a measure of available energy. The data were retrieved from continuous raster-based climatic maps with a resolution of 100 × 100 m (1991–2008) provided by the Meteorological Observatory of Friuli Venezia Giulia (OSMER).
For variables of human-impact, we quantified population density for each cell (POP) using the dasymetric grid of population density disaggregated with Corine Land Cover and point survey data, as described in
The composition of the landscape was described by the distribution of natural and semi-natural habitats in each cell, excluding agricultural and artificial habitats used as a proxy of human disturbance. Percentages of habitats were calculated from the CORINE biotopes map (1:50 000) of the study region (Servizio Valutazione Impatto Ambientale - Direzione Centrale Ambiente e Lavori Pubblici, Trieste, Italy; available at http://www.regione.fvg.it/ ). A standard classification of European habitats from the European Nature Information System (EUNIS; Davies et al., 2004, available at http://eunis.eea.europa.eu/habitats.jsp ) was chosen as a convenient platform for evaluating biological invasions in Europe (
Statistical analyses were performed with all alien plants occurring in the study region and within separate groups of alien species defined by residence time. First, the relative roles of climatic, human, and landscape variables on the observed variation in species richness were assessed using ordinary least squares (OLS) regression. Given that we had no a priori hypotheses supporting interactions between the considered explanatory variables, we did not include any interaction terms in our model selection procedure. We performed a backward manual deletion procedure starting from the full model (P < 0.05). We standardised the response variables (i.e. species richness) to zero mean and unit standard deviation to make the parameter estimates comparable. To produce a set of non-negative standardised variables, a constant value of three was added to all values.
Second, we evaluated the robustness of the standardised regression coefficients of our non-spatial OLS models by comparing them with those of spatial models generated with spatial eigenvector mapping (SEVM) techniques (
Third, we used hierarchical partitioning (HP) (
Finally, we conducted partial regressions to partition the variation explained by environmental (backward-selected climatic, human-impact, and landscape variables) and spatial variables into independent and covarying components. The total explained variation in species richness was partitioned into three components (
Range sizes decreased significantly from older to intermediate to recent neophytes (Fig. 1b, one-way ANOVA, F = 7.73, P < 0.001).
The multiple regression models showed several differences in the significance and rank of the standardised regression coefficients among separate groups of alien species defined by residence time (Table 1). For older neophytes, the model retained temperature and population density. For intermediate and recent neophytes, the models included temperature, population density, and thermophilous habitats. Temperature was the most important factor and was positively correlated with alien species richness in all the models. Positive partial coefficients were also found for the regression coefficients of the population density, but they were always substantially lower than those of temperature, especially for older neophytes. Thermophilous habitats had a weaker positive relationship with alien species in the models of intermediate and recent neophytes, suggesting that alien species richness is higher in dry and warm conditions.
Minimum adequate models for the relationships between alien species richness and the predictors (TEMP, annual mean temperature; POP, population density; TERM, thermophilous habitats) in the in the Friuli Venezia-Giulia region (Southern Alps) tested in the ordinary least squares (OLS) multiple regression models and in the spatial models generated with spatial eigenvector mapping (SEVM) techniques. The models are for (a) all neophyte plants occuring in the study region and within separate groups of neophytes defined by minimum residence time (MRT): (b) older neophytes (MRT = 200-500 yr), (c) intermediate neophytes (MRT = 100-200 yr), and (d) recent neophytes (MRT = <100 yr). Standardized regression coefficients (β) are presented.
| OLS | SEVM | |||||||
|---|---|---|---|---|---|---|---|---|
| β | t | P | R2 | β | t | P | R2 | |
| (a) All neophytes | 0.82 | 0.85 | ||||||
| TEMP | 0.558 | 7.895 | <0.001 | 0.815 | 3.543 | 0.001 | ||
| POP | 0.444 | 6.293 | <0.001 | 0.406 | 4.724 | <0.001 | ||
| TERM | 0.209 | 3.610 | <0.001 | 0.207 | 2.740 | 0.009 | ||
| (b) Older neophytes | 0.66 | 0.82 | ||||||
| TEMP | 0.685 | 7.033 | <0.001 | 0.705 | 4.735 | <0.001 | ||
| POP | 0.196 | 2.012 | 0.049 | 0.395 | 4.747 | <0.001 | ||
| (c) Intermediate neophytes | 0.81 | 0.90 | ||||||
| TEMP | 0.557 | 7.710 | <0.001 | 0.812 | 3.987 | <0.001 | ||
| POP | 0.447 | 6.187 | <0.001 | 0.319 | 4.329 | <0.001 | ||
| TERM | 0.177 | 2.995 | 0.004 | 0.028 | 0.390 | 0.699 | ||
| (d) Recent neophytes | 0.72 | 0.84 | ||||||
| TEMP | 0.471 | 5.343 | <0.001 | 0.246 | 1.478 | 0.147 | ||
| POP | 0.461 | 5.227 | <0.001 | 0.356 | 4.347 | 0.001 | ||
| TERM | 0.237 | 3.283 | 0.002 | 0.137 | 1.828 | 0.075 |
Including spatial filters in the model increased the amount of variance able to be explained, while the rank of the standardised regression coefficients of the variables was similar to OLS in all but the models for recent neophytes (Table 1). The effects of spatial filters on the P-values of the regression coefficients, however, were more important. Temperature was significant in the OLS model of recent neophytes but insignificant in the SEVM model and the thermophilous habitats in the models of intermediate and recent neophytes. Moreover, the standardised regression coefficients of predictors showed strong differences between the OLS and SEVM models. Temperature (e.g. for all and intermediate neophytes) and population density (e.g. for older neophytes) particularly had strong shifts in coefficients between the OLS and SEVM analyses (Table 1).
The results of hierarchical partitioning generally reflected those yielded by the regression models but produced slightly different results concerning the relative importance of some variables (Fig. 2a). The variable ranking indicated that both temperature and alpine habitats, followed by population density and precipitation, were the best predictors, with the highest independent effect on the number of alien species for older neophytes. For intermediate and recent neophytes, population density showed the largest independent effects followed by temperature. Considering the three sets of environmental variables (Fig. 2b), there was a reduction in the influence of climate and landscape, and an increase in the influence of human-impact, from the older neophytes to the recent neophytes (from 49% to 35% for climate, from 34% to 29% for landscape, and from 17% to 36% for human-impact).
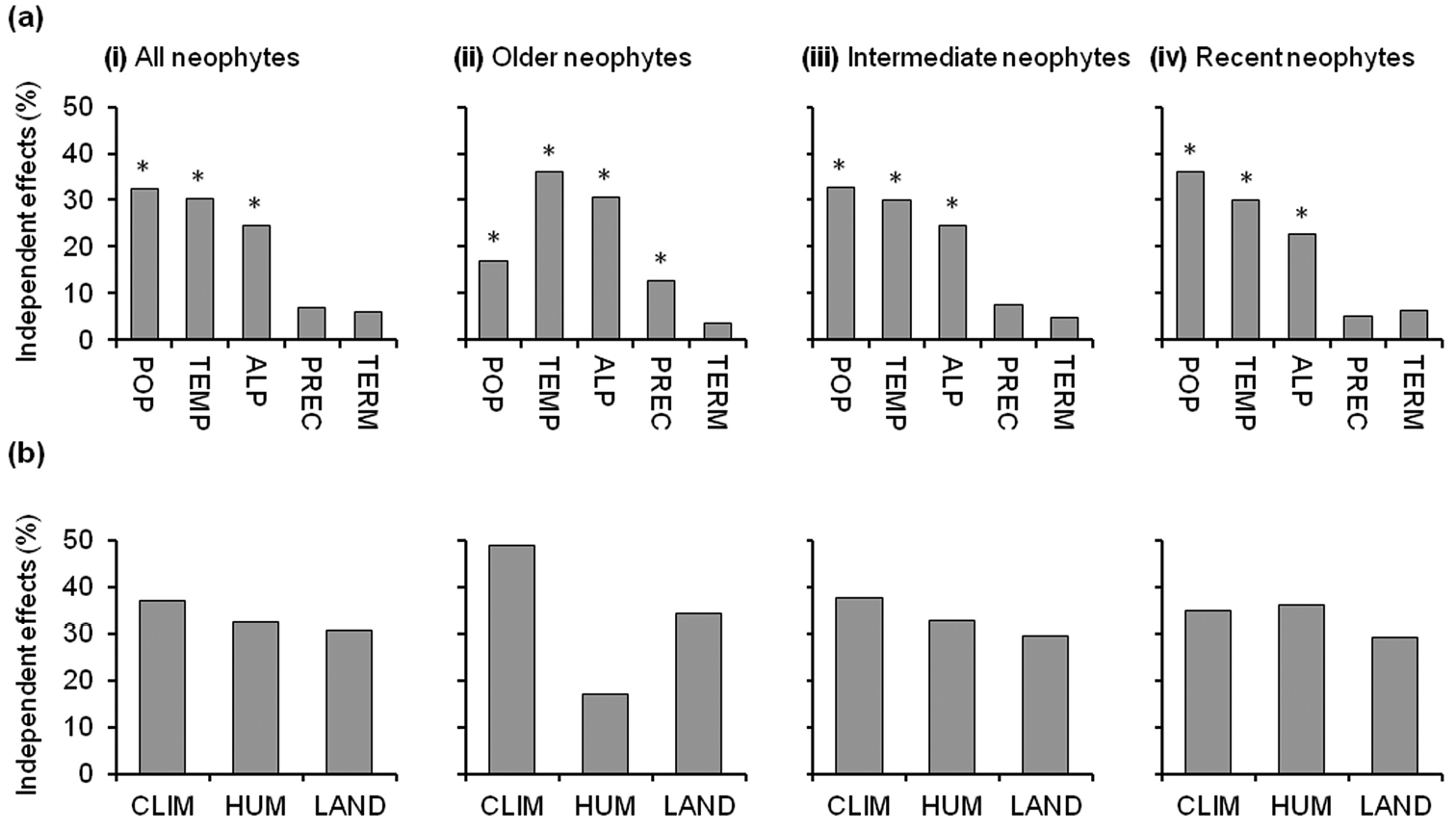
The relative independent effects of (a) environmental variables and (b) environmental variable sets (CLIM-climate, HUM-human-impact, and LAND-landscape) on species richness of (i) all neophytes, (ii) older neophytes, (iii) intermediate neophytes, and (iv) recent neophytes. Variables are ranked by the size of the independent effect in model (i) (i.e. all neophytes). Asterisks indicate statistical significance (P < 0.05) of the independent effect of each variable based on randomization tests (n = 200). POP, population density; TEMP, annual mean temperature; ALP, alpine habitats; PREC, annual precipitation; TERM, thermophilous habitats. Climate includes temperature and precipitation; human-impact includes population density; and landscape includes alpine and thermophilous habitats.
Partial regression analyses showed that most of the variation explained by the models is attributable to spatially structured environmental variation (E∩S, c.70-80%), while environment (E) and space (S) each had small independent effects (c.10-20%) (Fig. 3). The independent environmental effect (E), however, decreased and conversely the independent spatial effect (S) increased, from the older neophytes to the recent neophytes (from 14% to 9% for environment and from 11% to 18% for space).
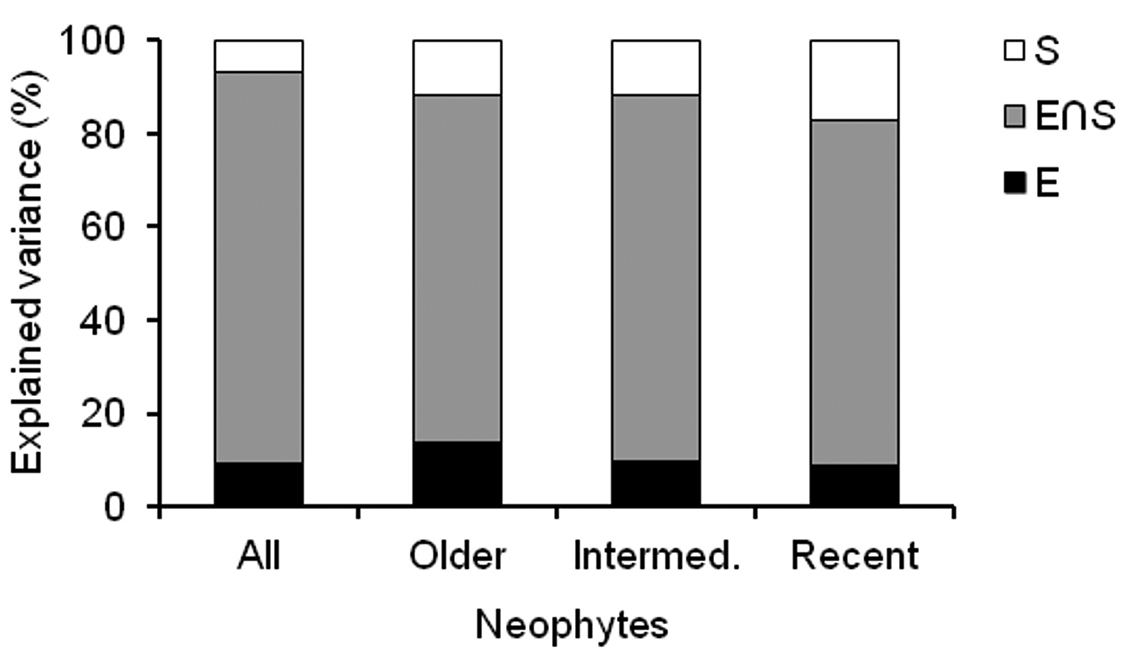
Variation partitioning of alien species richness per cell into environment and space for all neophytes, older neophytes, intermediate neophytes, and recent neophytes. The total variation explained was split into nonspatial environmental variation (E), spatially structured environmental variation (E∩S), and spatial structure not explained by the environmental variables (S). Results are expressed as part of the total explained variation for each component (i.e. values add up to 100%).
The observed tendency for the sizes of the ranges of alien species to increase with minimum residence time in the Friuli Venezia Giulia region is consistent with patterns reported elsewhere in the world (
This study also contributes to an understanding of how environmental-filtering (
Incorporating spatial filters in the models highlights several differences in the parameter estimates and shifts in coefficients between non-spatial and spatial regressions. These effects have also been observed in other studies (e.g.
Environmental variables (E + E∩S) account for c. 80-90% of the explained variation in alien species richness, although 70–75% of this variation is spatially-structured (E∩S). The shared variation between explanatory variables and spatial descriptors is produced by induced spatial dependence (
Specifically, we found strong differences in the importance of temperature and human disturbance on patterns of species richness in the three residence time groups. The association of alien species richness with temperature is stronger for groups with species having a longer residence time, which declines to become not significant for more recently arrived species, as shown in the spatial models. Furthermore, hierarchical partitioning shows a reduction in the influence of climate, from the older neophytes to the recent neophytes These results confirm the hypothesis that climate filtering (
Landscape variables instead show a trend similar to that observed for climate, with a reduction in the influence from the older neophytes to the recent neophytes. This trend could be due to the covariation among climatic and landscape variables. We particularly found a strong negative correlation between temperature and alpine habitats (r = -0.97; P < 0.001) and a meaningful negative correlation between precipitation and thermophilous habitats (r = -0.56; P < 0.001) (Appendix S2).
Role of spatial patternsThe shared variation between spatial descriptors and environmental variables is likely acting at broader spatial scales, accounting for environmental heterogeneity (e.g. climate and human pressures) and induced spatial dependence, whereas dispersal and biotic interactions such as competition, mortality, and social organisation are likely acting at finer spatial scales (
This study confirms the importance of considering residence time when studying spatial patterns of alien species richness and identifies residence time as a pivotal factor in the current distribution of alien species, i.e. climatic factors being most important for species with a longer residence time and factors related to human populations and habitat identity for species with shorter residence time. Our results also contribute to a better understanding of the influence of climate and human pressure on alien species richness and how these drivers shift their influences during the process of invasion. Additionally, the inclusion of spatial descriptors is important for explaining patterns of alien species and for unravelling the role of spatial autocorrelation generated by biotic processes.
We are grateful to Marisa Vidali for help in data collection and OSMER for providing climatic data. We thank William Blackhall for improving the English. We are grateful to two anonymous referees and the editor for the insightful comments that improved early drafts of the manuscript.
List of all predictors considered.
| Variable names and explanation | Unit | Mean ± SD | Min | Max | ||
|---|---|---|---|---|---|---|
| Climate | ||||||
| PREC | Annual precipitation | mm | 1551 ± 365 | 976 | 2465 | |
| TEMP | Annual mean temperature | °C | 10.18 ± 2.86 | 4.75 | 13.68 | |
| Human-impact | ||||||
| POP | Population density | per km2 | 126.5 ± 158.5 | 4.10 | 831.80 | |
| ROAD | Road length | km | 99.37 ± 61.90 | 3.88 | 258.70 | |
| URB | Total area covered by built-up elements | % | 7.30 ± 7.31 | 0.07 | 32.45 | |
| AGR | Area covered by agricultural area | % | 34.54 ± 34.06 | 0.00 | 89.55 | |
| Land-use | ||||||
| A2 | Littoral sediment | % | 1.73 ± 7.91 | 0.00 | 50.17 | |
| B1 | Coastal dunes and sandy shores | % | 0.01 ± 0.08 | 0.00 | 0.57 | |
| C1 | Surface standing waters | % | 0.13 ± 0.28 | 0.00 | 1.52 | |
| C2 | Surface running waters | % | 0.25 ± 0.45 | 0.00 | 1.97 | |
| C3 | Littoral zone of inland surface waterbodies | % | 1.65 ± 2.03 | 0.00 | 8.60 | |
| D2 | Valley mires, poor fens and transition mires | % | 0.03 ± 0.11 | 0.00 | 0.53 | |
| D5 | Sedge and reedbeds | % | 0.12 ± 0.61 | 0.00 | 4.26 | |
| E1 | Dry grasslands | % | 2.20 ± 4.35 | 0.00 | 21.73 | |
| E2 | Mesic grasslands | % | 1.60 ± 1.63 | 0.00 | 8.77 | |
| E3 | Seasonally wet and wet grasslands | % | 0.01 ± 0.03 | 0.00 | 0.17 | |
| E4.3 | Acid alpine and subalpine grassland | % | 0.64 ± 1.74 | 0.00 | 10.36 | |
| E4.4 | Calcareous alpine and subalpine grassland | % | 1.81 ± 2.68 | 0.00 | 10.14 | |
| E4.5 | Alpine and subalpine enriched grassland | % | 0.15 ± 0.42 | 0.00 | 1.86 | |
| F2 | Arctic, alpine and subalpine scrub | % | 4.47 ± 7.15 | 0.00 | 31.88 | |
| F3 | Temperate and mediterranean-montane scrub | % | 1.75 ± 3.49 | 0.01 | 21.39 | |
| F9 | Riverine and fen scrubs | % | 0.12 ± 0.27 | 0.00 | 1.47 | |
| G1.1 | Riparian and gallery woodland | % | 0.67 ± 0.97 | 0.00 | 3.37 | |
| G1.3 | Mediterranean riparian woodland | % | 0.00 ± 0.01 | 0.00 | 0.10 | |
| G1.4 | Broadleaved swamp woodland not on acid peat | % | 0.08 ± 0.22 | 0.00 | 1.05 | |
| G1.6 | [Fagus] woodland | % | 13.69 ± 16.63 | 0.00 | 55.24 | |
| G1.7 | Thermophilous deciduous woodland | % | 6.53 ± 9.51 | 0.00 | 46.33 | |
| G1.8 | Acidophilous [Quercus]-dominated woodland | % | 2.31 ± 5.66 | 0.00 | 27.32 | |
| G1.A | Meso- and eutrophic [Quercus] and related woodland | % | 1.78 ± 4.00 | 0.00 | 20.71 | |
| G3.1 | [Abies] and [Picea] woodland | % | 7.59 ± 15.17 | 0.00 | 55.17 | |
| G3.2 | Alpine [Larix] - [Pinus cembra] woodland | % | 1.42 ± 2.33 | 0.00 | 8.57 | |
| G3.5 | [Pinus nigra] woodland | % | 3.88 ± 6.47 | 0.00 | 23.82 | |
| H2 | Screes | % | 0.75 ± 1.31 | 0.00 | 6.84 | |
| H3 | Inland cliffs, rock pavements and outcrops | % | 1.37 ± 2.42 | 0.00 | 9.97 | |
| H4 | Snow or ice-dominated habitats | % | 0.02 ± 0.15 | 0.00 | 1.13 | |
Pearson correlations between explanatory variables (TEMP, annual mean temperature; PREC, annual precipitation; POP, population density; ROAD, road length; URB, total area covered by built-up elements; AGR, area covered by agricultural area; ALP, alpine habitats; TERM, thermophilous habitats).
| TEMP | PREC | POP | ROAD | URB | AGR | ALP | |
|---|---|---|---|---|---|---|---|
| PREC | -0.563 | ||||||
| POP | 0.574 | -0.412 | |||||
| ROAD | 0.721 | -0.325 | 0.744 | ||||
| URB | 0.711 | -0.505 | 0.916 | 0.827 | |||
| AGR | 0.843 | -0.699 | 0.498 | 0.651 | 0.724 | ||
| ALP | -0.971 | 0.482 | -0.512 | -0.654 | -0.631 | -0.754 | |
| TERM | 0.045 | -0.556 | 0.004 | -0.079 | 0.088 | 0.306 | 0.000 |
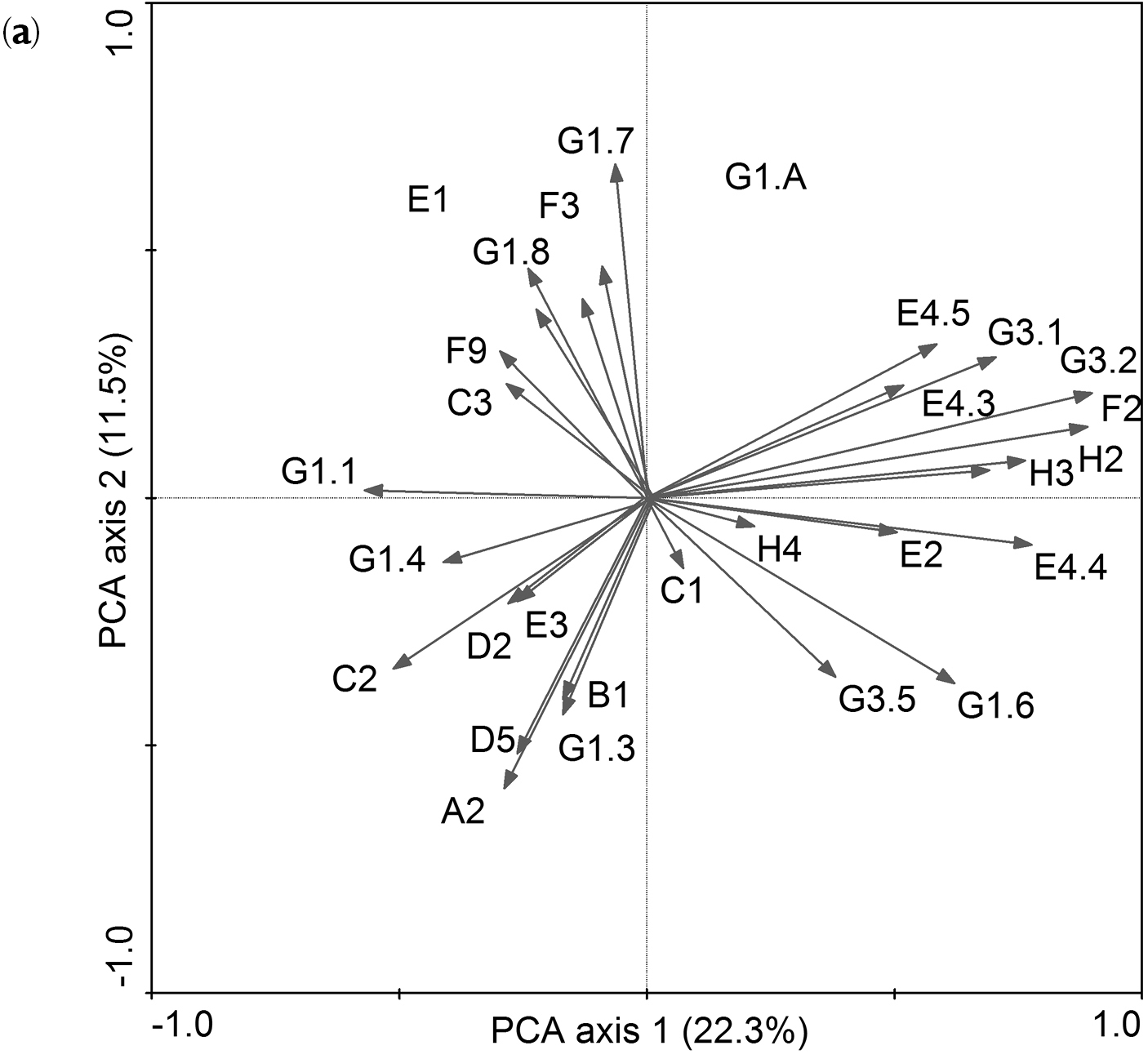
(a) Principal components analysis (PCA) diagram of 29 habitat classes, for codes of habitat classes please see Appendix S1
(b) Eigenvector scores of 29 habitat classes in two main PCA axes. Values are ranked in order of absolute magnitude along PCA 1. The five highest absolute eigenvector scores for each PCA axis are indicated in bold. Values in parentheses indicate variance accounted for by each axis.
| Habitat | PCA1 - ALP (22.3%) |
PCA2 - TERM (11.5%) |
|
|---|---|---|---|
| G3.2 | Alpine [Larix] - [Pinus cembra] woodland | 0.899 | -0.211 |
| F2 | Arctic, alpine and subalpine scrub | 0.890 | -0.143 |
| E4.4 | Calcareous alpine and subalpine grassland | 0.777 | 0.094 |
| H2 | Screes | 0.764 | -0.075 |
| G3.1 | [Abies] and [Picea] woodland | 0.704 | -0.284 |
| H3 | Inland cliffs, rock pavements and outcrops | 0.691 | -0.056 |
| G1.6 | [Fagus] woodland | 0.621 | 0.374 |
| E4.5 | Alpine and subalpine enriched grassland | 0.585 | -0.311 |
| G1.1 | Riparian and gallery woodland | -0.570 | -0.015 |
| E4.3 | Acid alpine and subalpine grassland | 0.518 | -0.227 |
| E2 | Mesic grasslands | 0.505 | 0.070 |
| C2 | Surface running waters | -0.492 | -0.349 |
| G3.5 | [Pinus nigra] woodland | 0.381 | 0.361 |
| G1.4 | Broadleaved swamp woodland not on acid peat | -0.322 | -0.095 |
| A2 | Littoral sediment | -0.282 | -0.644 |
| E3 | Seasonally wet and wet grasslands | -0.268 | -0.200 |
| D5 | Sedge and reedbeds | -0.262 | -0.600 |
| E1 | Dry grasslands | -0.260 | 0.494 |
| D2 | Valley mires, poor fens and transition mires | -0.249 | -0.188 |
| F9 | Riverine and fen scrubs | -0.238 | 0.246 |
| H4 | Snow or ice-dominated habitats | 0.217 | 0.058 |
| C3 | Littoral zone of inland surface waterbodies | -0.195 | 0.187 |
| B1 | Coastal dunes and sandy shores | -0.189 | -0.462 |
| G1.A | Meso- and eutrophic [Quercus] and related woodland | -0.187 | 0.330 |
| G1.3 | Mediterranean riparian woodland | -0.182 | -0.464 |
| G1.8 | Acidophilous [Quercus]-dominated woodland | -0.138 | 0.421 |
| F3 | Temperate and mediterranean-montane scrub | -0.085 | 0.484 |
| C1 | Surface standing waters | 0.073 | 0.142 |
| G1.7 | Thermophilous deciduous woodland | -0.040 | 0.672 |