NeoBiota 18: 151–156, doi: 10.3897/neobiota.18.5288
Could natural selection change the geographic range limits of light brown apple moth (Lepidoptera, Tortricidae) in North America?
Amy C. Morey 1, Robert C. Venette 2, William D. Hutchison 1
1 Department of Entomology, University of Minnesota, St. Paul, MN, USA
2 Northern Research Station, USDA Forest Service, St. Paul, MN 5518, USA
Citation: Morey AC, Venette RC, Hutchison WD (2013) Could natural selection change the geographic range limits of light brown apple moth (Lepidoptera, Tortricidae) in North America? In: Kriticos DJ, Venette RC (Eds) Advancing risk assessment models to address climate change, economics and uncertainty. NeoBiota 18: 151–156. doi: 10.3897/neobiota.18.5288
We artificially selected for increased freeze tolerance in the invasive light brown apple moth. Our results suggest that, by not accounting for adaptation to cold, current models of potential geographic distributions could underestimate the areas at risk of exposure to this species.
Forecasting future distributions of invasive insects is important for many management and regulatory decisions. However, numerous challenges exist in creating accurate, biologically relevant models and maps that are meaningful over time (Venette et al. 2010). In particular, no models currently account for the potential of an invasive species to adapt to a new environment. In fact, demographic models in invasion biology commonly treat species as “homogenous immutable entities” (Lee 2002). For invasions by alien species in North America, adaptation to cold temperature may be especially important at northern latitudes or high elevations; cold often prevents species from surviving year round (Huey 2010, Venette 2013). For example, cold is likely to constrain the future distribution of the light brown apple moth, Epiphyas postvittana (Walker), a recent insect invader to North America. No model for Epiphyas postvittana currently accounts for the possibility of evolution of increased cold tolerance. The objective of this study was to determine if it was possible to artificially select for increased cold tolerance in this species, and if so, to begin exploring the subsequent geographical repercussions. Epiphyas postvittana is considered to be predominately freeze intolerant during its purported overwintering stage, the late instar larva (Burgi and Mills 2010), but preliminary data suggests that a small proportion of the population may also be freeze tolerant (Venette unpublished data). This phenomenon could be considered at least “partial freeze tolerance” (Sinclair 1999), and enabled freezing to act as a strong selection pressure for enhanced cold tolerance in our study.
Epiphyas postvittana eggs were obtained from USDA-APHIS (permit P526P-11-03713). All subsequent rearing and experimentation was conducted in a Biosafety Level 2 Containment Facility in St. Paul, MN. Eggs were held at 23°C, 60% RH, and resulting neonates were reared on artificial bean diet until late instars. We cooled 4-6th instars (verified through head capsule measurement; Danthanarayana 1975) individually inside gelatin capsules at ~1°C/min to their supercooling point inside a 80°C freezer (modified from Carrillo et al. 2004). Once freezing occurred, larvae were immediately returned to 23°C and given fresh diet. Mortality was measured as failure to eclose. Surviving moths were randomly mated in 0.47L (16 fluid oz) containers with 1-3 individuals of each sex. Randomly selected offspring were subsequently reared and supercooled as previously described. This procedure was repeated for nine generations. A minimum of 102 larvae were supercooled in each generation. A control population was maintained simultaneously and identically, save exposure to freezing.
Survival following freezing after nine generations was compared between the selected and control populations using non-parametric cumulative incidence functions (CIF) in SAS 9.3 (SAS Institute, Cary, N.C.) to address competing risk (Satagopan et al. 2004). The competing risk was any individual that froze and survived. These estimates were then used to calculate the temperature required to kill 50% (LT50) of each population.
We used NAPPFAST (Magarey et al. 2009) to map where temperatures might fall below the LT50 of the selected or control populations. For each 10 x 10km grid cell, NAPPFAST calculated the proportion of the last 10 years in which the lowest temperature of the year was colder than the LT50 for each population. We ran the model with 3-D interpolated climate data.
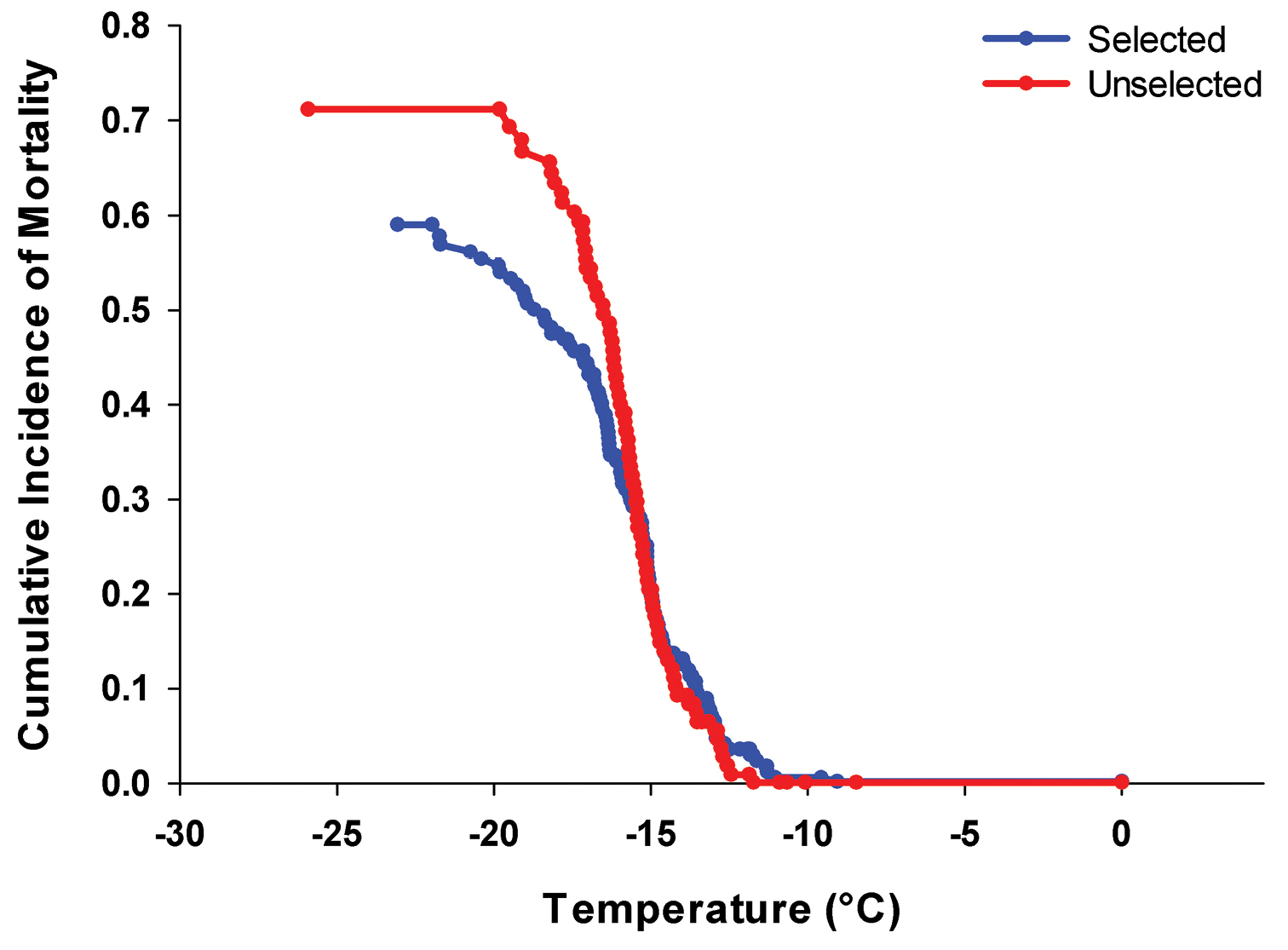
After only nine generations of selection, the probability of survival following freezing was significantly (α=0.1) greater for the selected population than the unselected control (Fig. 1; P=0.078, df=1, χ2=3.11). The LT50 for the unselected and selected populations were estimated to be –16.5°C and –19.0°C, respectively.
Figure 1.
Cumulative incidence of mortality of Epiphyas postvittana late instars with (n=179) and without (n=109) nine generations of artificial selection for increased cold tolerance.
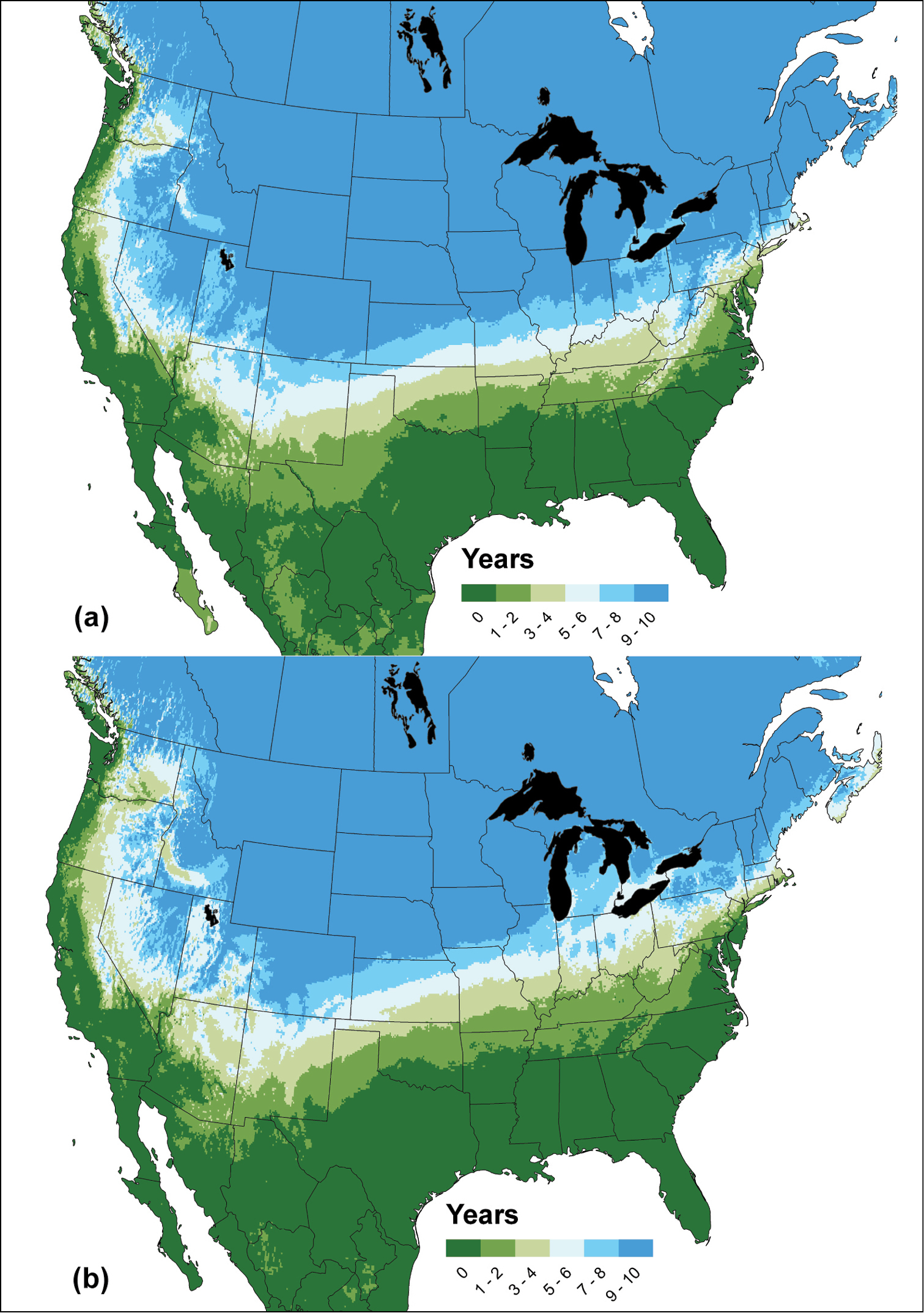
Figure 2 illustrates the geographic significance of a putative increase in cold tolerance for Epiphyas postvittana. Dark blue areas indicate the most dramatic effect of cold, where the LT50 was reached in 9-10 of the 10 years modeled. Cold was sufficient to exclude Epiphyas postvittana in many northern areas (e.g., Minnesota, the Dakotas, Wyoming, and much of Canada) using either the unselected (Fig. 2a) or selected (Fig. 2b) model. However, for other mid-western, eastern, and southern states, there was an overall reduction in the number of years where the LT50 was reached when using the selected population; the light blue to dark green colors shifted north. For example, the unselected model projected that nearly all of Michigan would reach the LT50 during 90–100% (dark blue) of the years modeled. In contrast, the selected model projected that most of the state would only reach the threshold between 50 to 80% of the modeled years (light blue and white). Similarly, western states (e.g., Nevada and Idaho) showed an eastward shift in the number of years that temperatures did not reach the LT50.
Figure 2.
The frequency of years (2003–2012) in which temperatures fell below the threshold required to cause 50% mortality (LT50) of Epiphyas postvittana in North America: a without selection, and b after nine generations of selection for increased freeze tolerance. Dark blue indicates where the LT50 was reached in 9–10 of the 10 years, dark green indicates where the LT50 was never reached during the 10-yr period.
Current risk maps that exist for Epiphyas postvittana (e.g., Fowler et al. 2009, Gutierrez et al. 2010, Lozier and Mills 2011), acknowledge the importance of cold in shaping this species’ potential U.S. range. However, the parameter(s) used to describe cold tolerance is/are assumed to be static. If natural selection follows a pattern similar to what our research suggests, within a relatively short period, current models may underestimate the risk of Epiphyas postvittana exposure in some areas in the future.
Uncertainty is inherent in pest risk models and contending with it is an ongoing area of research (Venette et al. 2010). Our study attempted to address the uncertainty related to the potential of a species to adapt to a cold environment and highlighted the geographic consequences if adaptation to cold is not considered. However, other sources of uncertainty still remain and are important future directions of this work. For example, the time of year may influence the effectiveness of selection on Epiphyas postvittana, assuming multi-voltinism and a randomly mating population. Selection is only likely to increase cold tolerance when there is a strong pressure (winter). But if there is any trade-off between increased cold tolerance and fitness (see Watson and Hoffman 1996, Huey 2010), cold adapted individuals may be selected against during months when selection pressure is reduced (summer). Similarly, particularly for a highly polyphagous insect like Epiphyas postvittana, host plant variability could also affect cold tolerance measures (e.g., Liu et al. 2009). Understanding the relationship between cold tolerance and these additional factors will undoubtedly continue to further enhance the accuracy and ultimate utility of pest risk mapping tools.
Acknowledgements
We thank Glen Fowler, Tracy Twine, and Tim Kurtti for comments on an earlier draft. We are also grateful for the assistance of Erica Nystrom, Laurel Mosca, Alex Sloane, and Kiley Friedrich in data collection, and to the MAES/MDA BSL-2 staff. This project was supported in part by a U.S. National Science Foundation (NSF)-Integrative Graduate Education and Research Traineeship Fellowship and the USDA Forest Service. We also thank NSF-Research Coordination Networks-FORECAST for funding A.C. Morey’s participation in the sixth annual meeting of the International Pest Risk Mapping Workgroup.
ReferencesBurgi LP, Mills NJ (2010) Cold tolerance of the overwintering larval instars of light brown apple moth
Epiphyas postvittana. Journal of Insect Physiology 56: 1645-1650. doi:
10.1016/j.jinsphys.2010.06.009Danthanarayana W (1975) The bionomics, distribution and host range of the light brown apple moth,
Epiphyas postvittana (Walk.) (Tortricidae). Aust. J. Zoology 23: 419-437. doi:
10.1071/ZO9750419Fowler G, Garrett L, Neeley A, Magarey R, Borchert D, Spears B (2009) Economic Analysis: Risk to U.S. apple, grape, orange, and pear production from the light brown apple moth, Epiphyas postvittana (Walker). USDAA-PHIS-PPQ-CPHST-PERAL, 28 pp.
http://www.aphis.usda.gov/plant_health/plant_pest_info/lba_moth/downloads/lbameconomicanalysis.pdfGutierrez AP, Mills NJ, Ponti L (2010) Limits to the potential distribution of light brown apple moth in Arizona–California based on climate suitability and host plant availability. Biological Invasions 12: 3319-3331. doi:
10.1007/s10530-010-9725-8Huey RB (2010) Evolutionary physiology of insect thermal adaptation to cold environments. In: Denlinger D, Lee R (Eds) Low temperature biology of insects. Cambridge Univ. Press, New York, 223-241.
Lozier JD, Mills NJ (2011) Predicting the potential invasive range of light brown apple moth (
Epiphyas postvittana) using biologically informed and correlative species distribution models. Biological Invasions 13: 2409-2421. doi:
10.1007/s10530-011-0052-5Liu Z, Gong P, Heckel DG, Wei W, Sun J, Li D (2009) Effects of larval host plants on over-wintering physiological dynamics and survival of the cotton bollworm,
Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae). Journal of Insect Physiology 55: 1-9. doi:
10.1016/j.jinsphys.2008.07.017Satagopan JM, Ben-Porat L, Berwick M, Robson M, Kutler D, Auerbach AD (2004) A note on competing risks in survival data analysis. British Journal of Cancer 91: 1229-1235. doi:
10.1038/sj.bjc.6602102Sinclair BJ (1999) Insect cold tolerance: How many kinds of frozen? European Journal of Entomology 96: 157–164.
Venette RC (2013) Incorporating climate change into pest risk models for forest pathogens: a role for cold stress in an era of global warming? In: Kriticos DJ, Venette RC (Eds) Advancing risk assessment models to address climate change, economics and uncertainty. NeoBiota 18: 131–150. doi:
10.3897/neobiota.18.4047Venette RC, Kriticos DJ, Magarey RD, Koch FH, Baker RHA, Worner SP, Gomez Raboteaux NN, McKenney DW, Dobesberger EJ, Yemshanov D, De Barro PJ, Hutchison WD, Fowler G, Kalaris TM, Pedlar J (2010) Pest risk maps for invasive alien species: A roadmap for improvement. BioScience 60: 349–362.
http://www.forestthreats.org/www/efetac4/efetac4/products/publications/Pest_risk_maps_for_invasive_alien_species.pdf/view doi:
10.1525/bio.2010.60.5.5