





(C) 2014 Rethabile Frangenie Motloung. This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Citation: Motloung RF, Robertson MP, Rouget M, Wilson JRU (2014) Forestry trial data can be used to evaluate climate-based species distribution models in predicting tree invasions. NeoBiota 20: 31–48. doi: 10.3897/neobiota.20.5778
Climate is frequently used to predict the outcome of species introductions based on the results from species distribution models (SDMs). However, despite the widespread use of SDMs for pre- and post-border risk assessments, data that can be used to validate predictions is often not available until after an invasion has occurred. Here we explore the potential for using historical forestry trials to assess the performance of climate-based SDMs. SDMs were parameterized based on the native range distribution of 36 Australian acacias, and predictions were compared against both the results of 150 years of government forestry trials, and current invasive distribution in southern Africa using true skill statistic, sensitivity and specificity. Classification tree analysis was used to evaluate why some Australian acacias failed in trials while others were successful. Predicted suitability was significantly related to the invaded range (sensitivity = 0.87) and success in forestry trials (sensitivity = 0.80), but forestry trial failures were under-predicted (specificity = 0.35). Notably, for forestry trials, the success in trials was greater for species invasive somewhere in the world. SDM predictions also indicate a considerable invasion potential of eight species that are currently naturalized but not yet widespread. Forestry trial data clearly provides a useful additional source of data to validate and refine SDMs in the context of risk assessment. Our study identified the climatic factors required for successful invasion of acacias, and accentuates the importance of integration of status elsewhere for risk assessment.
Species distribution models, model evaluation, Australian acacias, classification tree, forestry, alien trees, invasions, Southern African Plant Invaders Atlas
Predicting which species will escape from forestry plantations and become invasive remains a challenge in invasion biology (
Species distribution models (SDMs) have been widely used to predict invasions (
The use of SDMs in management of invasive species can be considerably improved by independent datasets to validate SDM predictions (
While alien trees and shrubs have been introduced in different parts of the world to fulfil a wide range of human needs (
The aims of this study are to evaluate species distribution models using both forestry trial data and invaded range data, and to investigate why some trials succeeded while others failed. To do this, we narrowed the taxonomic range to a well studied group, Australian acacias. Australian acacias are a good model group to understand plant invasions because Acacia is a speciose genus that contains many introduced and invasive species (
The number of government forestry trials, the number of successful trials; the number of records in SAPIA, the number of QDGCs occupied in South Africa, Lesotho and Swaziland; and status in southern Africa for the species explored in the study.
| Species | Total number of trials | Success | Number of records in SAPIA | QDGCs occupied in SA | Status in southern Africa |
|---|---|---|---|---|---|
| Acacia acuminata | 13 | 3 | 0 | 0 | Introduced |
| Acacia adunca | 1 | 0 | 2 | 2 | Naturalized |
| Acacia adsurgens | 2 | 2 | 0 | 0 | Introduced |
| Acacia aneura | 4 | 3 | 0 | 0 | Introduced |
| Acacia aulacorpa | 4 | 2 | 0 | 0 | Introduced |
| Acacia auriculiformis | 3 | 2 | 0 | 0 | Introduced |
| Acacia baileyana | 5 | 4 | 184 | 101 | Invasive |
| Acacia binervata | 1 | 1 | 0 | 0 | Introduced |
| Acacia cowleana | 2 | 1 | 0 | 0 | Introduced |
| Acacia crassicarpa | 2 | 1 | 0 | 0 | Introduced |
| Acacia cultriformis | 7 | 3 | 1 | 1 | Naturalized |
| Acacia cyclops | 2 | 2 | 1282 | 172 | Invasive |
| Acacia dealbata | 13 | 12 | 1667 | 299 | Invasive |
| Acacia decurrens | 5 | 2 | 341 | 124 | Invasive |
| Acacia elata | 9 | 3 | 99 | 48 | Invasive |
| Acacia falciformis | 4 | 2 | 0 | 0 | Introduced |
| Acacia fimbriata | 1 | 1 | 1 | 1 | Naturalized |
| Acacia holosericea | 4 | 1 | 1 | 0 | Introduced |
| Acacia implexa | 0 | 0 | 3 | 3 | Invasive |
| Acacia leptocarpa | 2 | 2 | 0 | 0 | Introduced |
| Acacia ligulata | 1 | 1 | 0 | 0 | Introduced |
| Acacia longifolia | 5 | 4 | 446 | 97 | Invasive |
| Acacia mangium | 3 | 1 | 0 | 0 | Introduced |
| Acacia mearnsii | 13 | 10 | 4313 | 462 | Invasive |
| Acacia melanoxylon | 28 | 20 | 678 | 167 | Invasive |
| Acacia paradoxa | 1 | 1 | 4 | 2 | Invasive |
| Acacia pendula | 6 | 3 | 0 | 0 | Introduced |
| Acacia podalyriifolia | 3 | 2 | 159 | 78 | Invasive |
| Acacia prominens | 2 | 2 | 0 | 0 | Introduced |
| Acacia pycnantha | 9 | 8 | 182 | 38 | Invasive |
| Acacia retinodes | 2 | 2 | 0 | 0 | Introduced |
| Acacia rubida | 1 | 1 | 0 | 0 | Introduced |
| Acacia saligna | 8 | 5 | 1302 | 164 | Invasive |
| Acacia schinoides | 1 | 1 | 0 | 0 | Introduced |
| Acacia stricta | 0 | 0 | 6 | 6 | Invasive |
| Acacia viscidula | 1 | 1 | 1 | 1 | Naturalized |
| Total | 168 | 109 | 10672 | 1766 |
First we used data on the outcome of forestry trials and data on invasive plant distributions to evaluate SDMs. We then assessed why some forestry trials succeeded but others failed. Lastly, we determine which introduced species have a large potentially suitable but not currently occupied range, i.e. which species have a high invasion debt (Essl et al. 2011).
Species distribution models for Australian acacias were calibrated using occurrence records from their native range (Australia) and then projected to southern Africa based on
Model calibration
Numerous approaches have been developed to model the potential distribution of invasive species with various degrees of success (
We obtained a dataset of occurrence records for the selected species (Table 1) from the Australian Virtual Herbarium (accessed 29th June 2010) with only one occurrence record retained per 10 minute grid cell for each species to reduce spatial sampling bias.
For each species we calculated the minimum and maximum values extracted from occurrence records for each predictor variable. These values were used to identify the range of values that each species could tolerate. For each species, we reclassified each of the four predictor variable maps into a map consisting of presence (value = 1) or absence (value = 0), using the minimum and maximum values calculated for that species. We then multiplied these presence/absence maps to generate a final map indicating the potential distribution of that species. The analysis was conducted using the R statistical software (v. 2.11,
Model evaluation
We evaluated the relationship between predicted distributions and actual distributions (forestry trials and SAPIA data) using several measures. Sensitivity is the proportion of observed presences predicted present and quantifies the omission error; specificity is the proportion of observed absences predicted absent and quantifies commission error (
Government forestry trials
In southern Africa, forestry trials for Australian acacias were conducted at 67 stations from the 1820s to the 1960s for 36 species (
Of the 168 forestry trial records obtained from this dataset, 129 records had precise geographic location and trial outcome and could be used to evaluate the SDMs (see Table 1). While subspecific taxa can occupy different climatic niches (
Southern African Plant Invaders Atlas (SAPIA)
This dataset contains records for over 700 naturalized and invasive species, with information on abundance, habitat preferences, time of introduction and distribution (
A classification tree approach (
Variables used to quantify why some forestry trials of Acacia species in southern Africa succeeded while others failed.
| Variable | Coded | Criteria | Justification | Data sources |
|---|---|---|---|---|
| Trial outcome | 0/1 | 0- failure (absence) 1-success (presence) |
|
|
| Biotic | Y/N | Did species fail because seeds were eaten by animals and birds? | Biotic factors have been indicated to influence survival that can lead to naturalization ( |
|
| Climatic suitability | 0/1 | 0-Climate not suitable 1-Climate suitable |
Climatic suitability is widely used to predict outcome of introductions McGregor et al. (2012). | Species distribution models |
| Climatic variables | Bio_1, bio_5, bio_6, bio_12 | Annual Mean temperature, Maximum Temperature of the Warmest Month, Minimum Temperature of the Coldest Month & Annual precipitation | Did trials succeed because of precipitation and temperature ranges? |
|
| Biome | Names | South African biomes | Were trials successful because of the biomes they were introduced to? |
|
| Status | Introduced, Naturalized or invasive (sensu |
Invasiveness elsewhere according to |
Invasiveness elsewhere is one of the best predictors of the outcome of species introductions |
|
The classification tree was constructed by repeatedly splitting data, defined by a simple rule based on a single explanatory variable at each split. At each split, the data were partitioned into two exclusive groups, each of which was as homogeneous as possible (
Occupancy of each species was calculated at the quarter degree grid cell (QDGC, 15 min. × 15 min.) level. The current distribution of Australian acacias in Lesotho, South Africa, and Swaziland was based on the number of QDGCs occupied as recorded in both the SAPIA and the forestry trial databases. Species potential distribution was estimated based on climatic suitability and range size as the percentage of southern Africa (South Africa, Lesotho and Swaziland) predicted suitable by SDMs.
The SDMs successfully predicted the outcome (success or failure) for 81 of the 129 forestry trials (63%) (overall TSS = 0.15, Table 3), with a high percentage of true presences predicted as present (sensitivity = 0.80) but a rather low percentage of true absences predicted as absent (specificity = 0.35). However, there was a higher proportion of false absences than false presences (Table 3). SDM evaluation varied between species, with five species having low sensitivity values and ten with low specificity values (Table 3).
Measures used to evaluate models using government forestry trials dataset in southern Africa; a-number of cells for which presence was correctly predicted by the model; b- number of cells for which the species was not found but the model predicted presence; c, number of cells for which the species was found and the model predicted absence; d, number of cells for which absence was correctly predicted by the model. Sensitivity (proportion of actual presences predicted as such); specificity (proportion of actual absences predicted as such); TSS [true skill statistic (sensitivity + specificity-1].
| Species | Number of trials | a | b | c | d | Sensitivity | Specificity | TSS |
| Acacia acuminata | 13 | 1 | 5 | 1 | 6 | 0.50 | 0.54 | 0.04 |
| Acacia aneura | 4 | 1 | 1 | 2 | 0 | 0.33 | 0.00 | -0.67 |
| Acacia baileyana | 5 | 4 | 1 | 0 | 0 | 1.00 | 0.00 | 0.00 |
| Acacia cultriformis | 5 | 0 | 2 | 1 | 2 | 0.00 | 0.50 | -0.50 |
| Acacia dealbata | 13 | 11 | 0 | 1 | 1 | 0.91 | 1.00 | 0.91 |
| Acacia decurrens | 7 | 2 | 4 | 0 | 1 | 1.00 | 0.20 | 0.20 |
| Acacia elata | 9 | 2 | 4 | 2 | 1 | 0.50 | 0.20 | -0.30 |
| Acacia falciformis | 4 | 2 | 2 | 0 | 0 | 1.00 | 0.00 | 0.00 |
| Acacia longifolia | 5 | 4 | 1 | 0 | 0 | 1.00 | 0.00 | 0.00 |
| Acacia mearnsii | 13 | 7 | 2 | 3 | 1 | 0.70 | 0.33 | 0.03 |
| Acacia melanoxylon | 28 | 20 | 8 | 0 | 0 | 1.00 | 0.00 | 0.00 |
| Acacia pendula | 6 | 1 | 1 | 2 | 2 | 0.33 | 0.67 | 0.00 |
| Acacia pycnantha | 9 | 7 | 1 | 1 | 0 | 0.88 | 0.00 | -0.20 |
| Acacia saligna | 8 | 2 | 0 | 3 | 3 | 0.40 | 1.00 | 0.40 |
| Overall | 129 | 64 | 32 | 16 | 17 | 0.80 | 0.35 | 0.15 |
Observed invasive ranges of Australian acacias in southern Africa are generally correctly predicted as suitable based on the SDMs (sensitivity = 0.87). However, the predictions mismatched 461 (13%) invasive range records (i.e. 13% of the records indicate that species invaded areas that are climatically unsuitable; Table 4). Most of the invasive species have the potential to substantially increase their ranges because more than half of southern Africa is predicted to be suitable.
Measures used to evaluate models using Southern African Plant Invaders Atlas; a-number of cells for which presence was correctly predicted by the model; c-number of cells for which the species was found but the model predicted absence.
| Species | Number of records | a | c | Sensitivity |
|---|---|---|---|---|
| Acacia baileyana | 86 | 82 | 4 | 0.95 |
| Acacia cyclops | 141 | 137 | 4 | 0.97 |
| Acacia dealbata | 717 | 707 | 10 | 0.99 |
| Acacia decurrens | 157 | 155 | 2 | 0.99 |
| Acacia elata | 20 | 17 | 3 | 0.85 |
| Acacia longifolia | 79 | 79 | 0 | 1.00 |
| Acacia mearnsii | 1916 | 1485 | 431 | 0.78 |
| Acacia melanoxylon | 163 | 163 | 0 | 1.00 |
| Acacia podalyriifolia | 56 | 51 | 5 | 0.91 |
| Acacia pycnantha | 83 | 82 | 1 | 0.99 |
| Acacia saligna | 53 | 52 | 1 | 0.98 |
| Overall | 3471 | 3010 | 461 | 0.87 |
Acacia dealbata and Acacia mearnsii are provided as visual examples (Fig. 1).
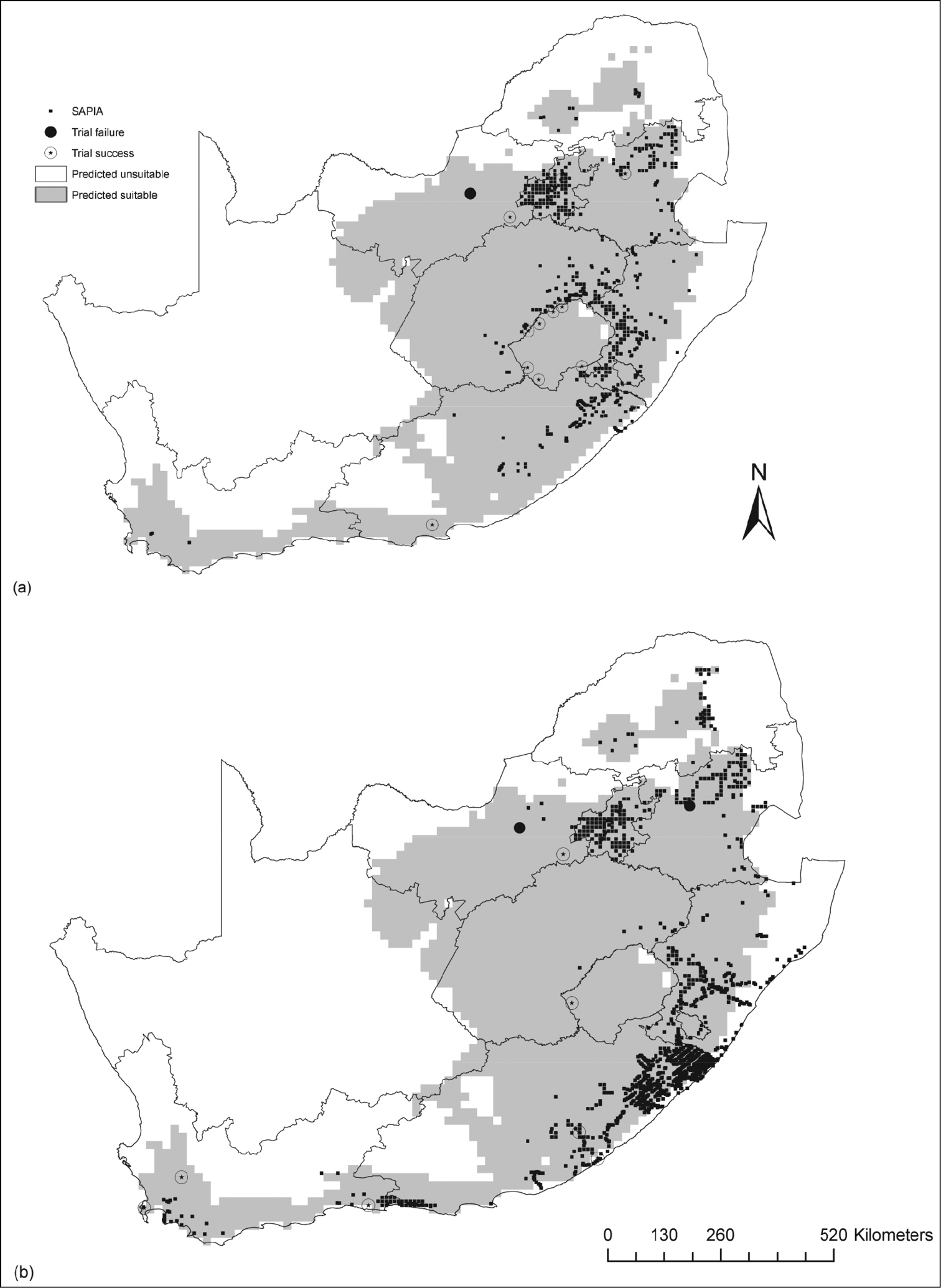
Current distribution [from both forestry trials (noted as trial failure or success) and SAPIA] and potential distribution for: a Acacia dealbata and b Acacia mearnsii in Lesotho, South Africa, and Swaziland.
Using climatic suitability alone, 81 trials out of 129 (model accuracy = 0.63) were correctly predicted (Table 3), while 87 (model accuracy = 0.67) records were correctly predicted by the criterion “invasive elsewhere” (Fig. 2: Node 2 and 3). Based on the classification tree (Fig. 2), three variables (invasive elsewhere, annual mean temperature, and mean temperature of the coldest month) together accurately predicted 72% of the forestry trial records (i.e. 93 trials). The classification tree analysis correctly predicted forestry trial outcome significantly (sensitivity = 086, specificity = 0.36 and TSS = 0.22).

Classification tree analysis of the probability that a species will succeed (blue part of the bar) or fail (red) in southern African forestry trials based on various variables (see Table 2: trial data and variables used). Each node shows the number of successes and failures in each class, the total number of cases (N) and a graphical presentation of the percentage of success and failure cases (horizontal bar). For each node the splitting criterion is written in caps on top of the nodes. Status is similar to invasiveness elsewhere and the two environmental variables: Bio_1 is annual mean temperature and Bio_6 is minimum temperature of the coldest month.
The wetter parts of the region were identified as most suitable for the analysed suite of Australian acacias. SDMs predicted that thirteen species can be widespread with 20 % of southern Africa predicted to be suitable [Acacia acuminata, Acacia adunca, Acacia aneura, Acacia cultriformis, Acacia falciformis, Acacia fimbriata, Acacia implexa, Acacia paradoxa, Acacia pendula, Acacia retinodes, Acacia rubida, Acacia stricta, and Acacia viscidula (Fig. 3)]. The remaining four species (Acacia mangium, Acacia prominens, Acacia schinoides and Acacia binervata), appear to have a potential of localized extensive spread (Fig. 3). All of these species have not yet reached the full extent of climatically suitable ranges (see Appendix – Fig. S1).
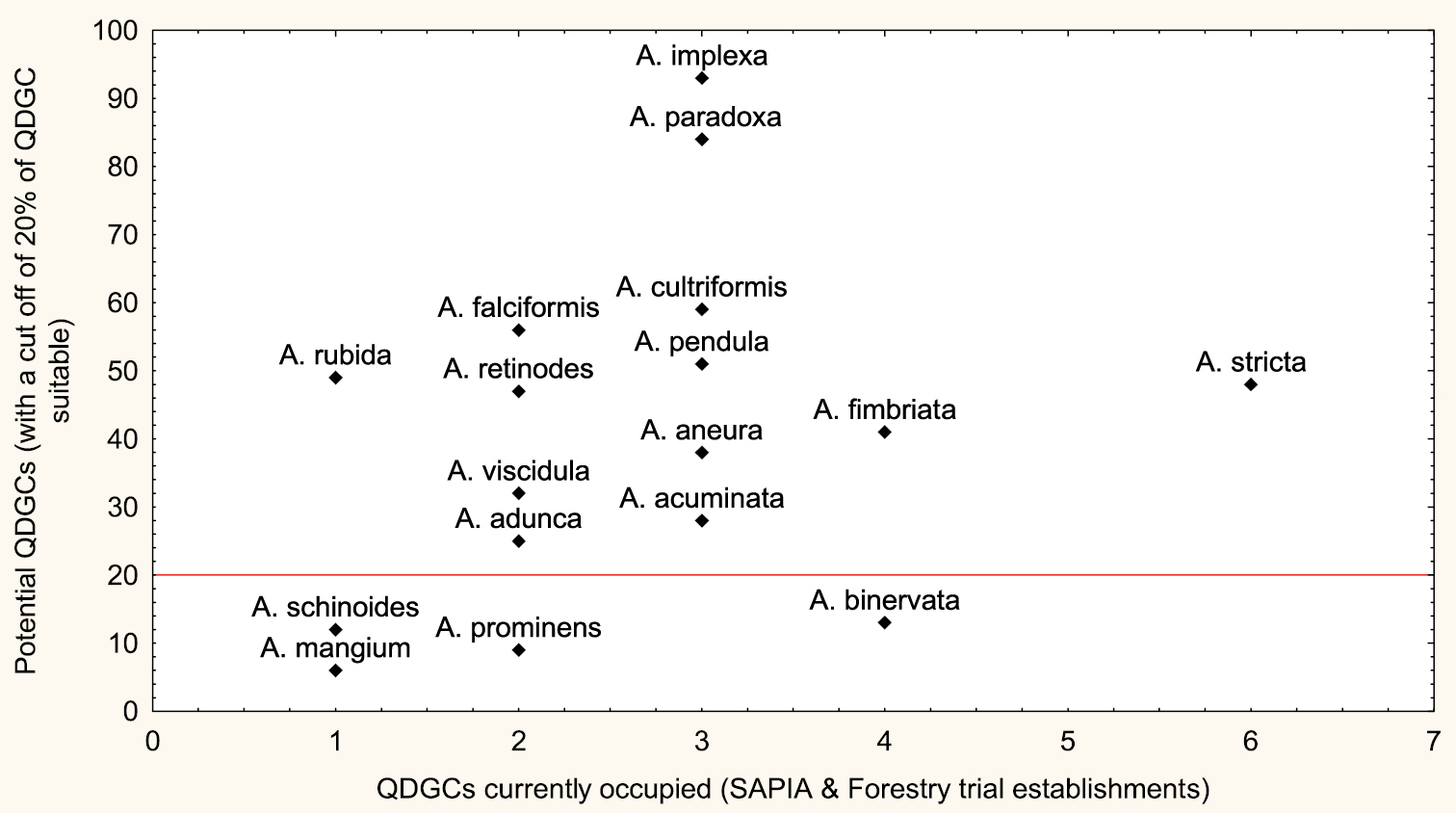
The relationship between potential and current distributions in Lesotho, South Africa, and Swaziland expressed as total number of QDGCs currently occupied from SAPIA and forestry trial successes (with a cut off of 20% of southern Africa QDGCs climatically suitable) for 17 species that are not yet widespread in southern Africa.
In this study, we found that forestry trial data provided useful data for SDM evaluation as it contains information on species that were repeatedly introduced in different localities within southern Africa with a clear indication of trial outcome. Trials were successful in areas that are predicted as climatically suitable by SDMs (sensitivity = 0.80; Table 3).
SDMs could not accurately predict the failures of species in trials (specificity = 0.35; Table 3) and this suggests that climatic suitability only provides a broad picture of where an introduced species might survive, but there are other factors that determine whether an introduced species will be successful like biotic interactions and evolutionary capacity to adapt (
SDMs correctly identified invaded ranges of Australian acacias in southern Africa. SDMs correctly predict regions of introduction but not the total invaded ranges (e.g. Acacia mearnsii, Fig. 1). Similarly, previous studies indicated that SDMs calibrated on native range records were able to predict region of introduction, but not the total invaded range, indicating a possible niche shift (
In line with other studies, we found that the success of species after introduction in the new ranges was greater for species recorded as invasive elsewhere (
Australian Acacia species that are not yet widespread are likely to spread to currently unoccupied climatically suitable ranges as SDMs predictions indicate that a large portion of Lesotho, South Africa and Swaziland is suitable for invasion by 13 of the 17 currently introduced or naturalized (Fig. 3). We believe this is a major invasion debt and not simply an over-prediction in the SDMs, because there is a strong correlation between extent of usage and invasive distributions for Australian acacias in South Africa (
The project was funded by the South African Department of Environment Affairs’ Working for Water (WfW) Programme. We are grateful to Lesley Henderson and R. J. Poynton for data on Australian acacias. Mathieu Rouget acknowledges support from the South African Research Chair Initiative of the Department of Science and Technology and National Research Foundation of South Africa.
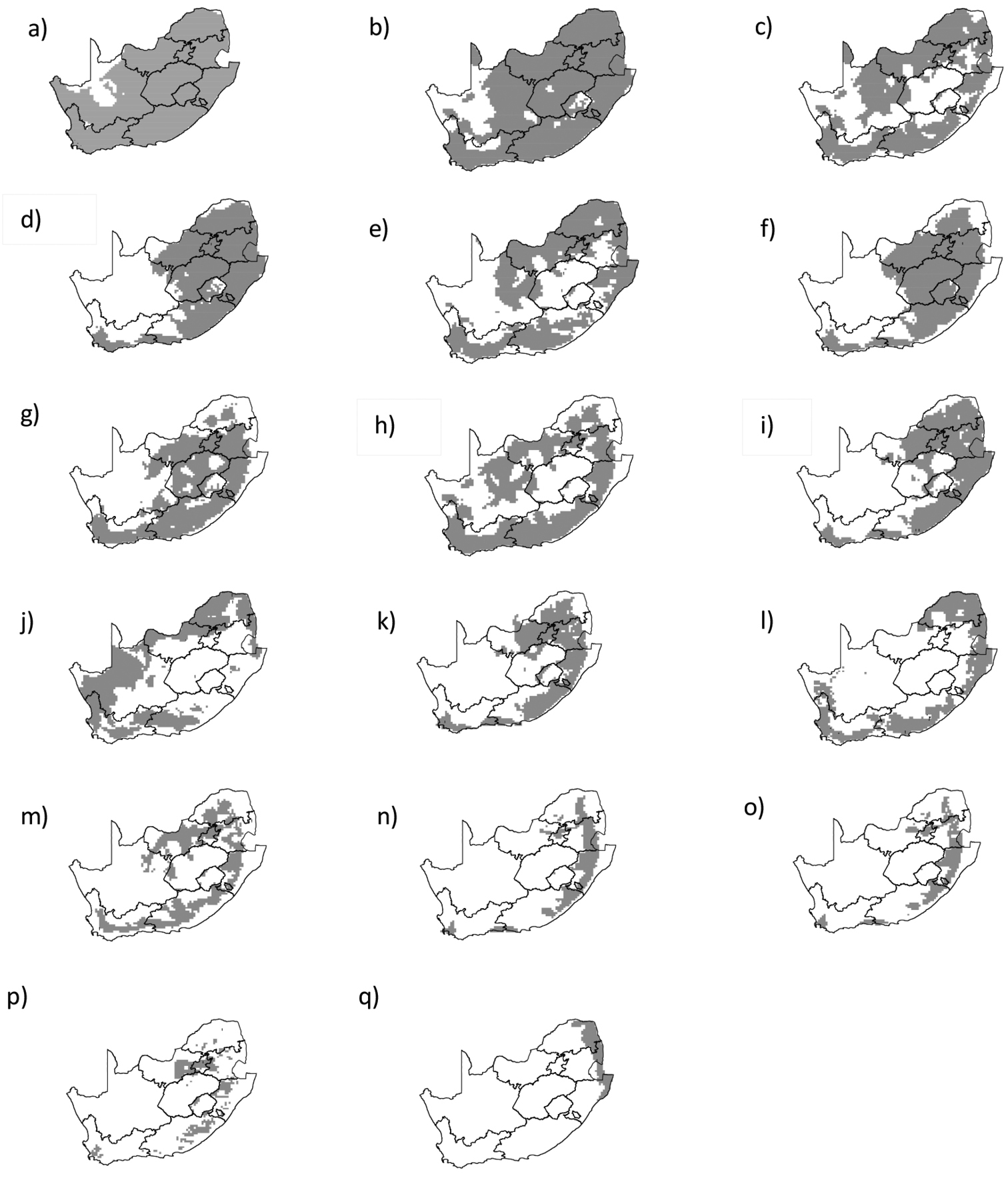
Current and potential distributions of sixteen species that are not widespread in southern Africa arranged on the basis of their suitable range size : a Acacia paradoxa b Acacia implexa c Acacia cultriformis d Acacia falciformis e Acacia pendula f Acacia rubida g Acacia stricta h Acacia retinodes i Acacia fimbriata j Acacia aneura k Acacia viscidula l Acacia acuminata m Acacia adunca n Acacia binervata o Acacia schinoides p Acacia prominens q Acacia mangium. The grey shading indicates areas that SDMs have identified as suitable by SDMs while the white shading indicates unsuitable areas.