Research Article |
|
Corresponding author: Pablo García-Díaz ( Garcia-DiazP@landcareresearch.co.nz ) Academic editor: Sven Bacher
© 2019 Pablo García-Díaz, Joshua V. Ross, Miquel Vall-llosera, Phillip Cassey.
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
García-Díaz P, Ross JV, Vall-llosera M, Cassey P (2019) Low detectability of alien reptiles can lead to biosecurity management failure: a case study from Christmas Island (Australia). NeoBiota 45: 75-92. https://doi.org/10.3897/neobiota.45.31009
|
Abstract
When we assume that contemporary management actions will be effective against the global rise of emerging alien species, we can develop management complacency, which leads to potentially disastrous outcomes for native biodiversity. Here, we propose the use of the probability of detection as a metric to assess the feasibility of management actions for alien species. We explore how detectability can influence the management of alien reptiles, a group of emergent alien vertebrates globally. We use a Rapid Biological Assessment method (time-limited transects) to estimate the probability of detection for alien reptiles present on Christmas Island (Australia). Across the five species studied, we found low probabilities of detection and poor explanatory capacity of the individual covariates included in our models. These findings indicate that management options to deal with alien reptiles are limited due to the potential high cost and low efficacy associated with low probabilities of detection. Strict preventive strategies, firmly espousing the principles of adaptiveness and precautionary policies, combined with early detection and biosecurity response activities are needed to address the emergence of alien reptiles. Our research was focussed on alien reptiles on islands, but the rise of new pools of alien species from all taxonomic realms across the world suggests that our conclusions may be applicable more generally. Further research is called for to explore the applicability of our conclusions and recommendations to other taxonomic groups and regions of the world.
Keywords
Anticipatory policy-making, Christmas Island, preventive decision-making, probability of detection, Rapid Biological Assessment, uncertainty
Introduction
The global emergence of a new pool of alien species may render existing management actions, ranging from rapid incursion response to the eradication of self-sustaining populations, ineffective (
Successful management of alien species depends on the capacity to anticipate the specific transport pathways, which move alien species, and on adapting management actions to address the associated novel risk. While substantial research effort has been invested in understanding the dynamics of changing pathways (
Rapid Biological Assessments (RBAs) are a widely used tool for conducting biodiversity inventories and monitoring, balancing reliability in sampling biological communities with time and resource constraints to conduct the surveys (
Reptiles are a notable group of emergent alien species responsible for serious environmental impacts in recipient regions worldwide, especially on islands (
We investigated a standardised RBA using the case study of the detection of alien reptiles on Christmas Island (Fig.

Methods
Study area and survey sites
Christmas Island is an Australian oceanic territory located in the Indian Ocean, 10°30’S 105°40’E (Fig.

Estimated probabilities of individual detection of five invasive alien reptiles on Christmas Island (Australia) during day time (top panel) and night time surveys (bottom panel). Estimates correspond to the probability of individual detection during a 10-minute survey across the 34 surveying sites. Each dot represents a realisation from 1000 simulations.
We identified 34 survey sites (Fig.
Goodness of fit (Bayesian p-values), summary statistics of the covariates (before standardisation; mean ± standard deviation, and range), and posterior coefficient estimates (mean ± standard error, and 95% Credible Intervals) of the abundance-detection models for five species of alien reptiles on Christmas Island.
| Covariate summary statistics (original units) | Common house gecko (Hemidactylus frenatus) | Stump-toed gecko (Gehyra mutilata) | Grass skink (Lygosoma bowringii) | Flowerpot snake (Indotyphlops braminus) | Wolf snake (Lycodon capucinus) | |
| Bayesian p-value | 0.51 | 0.49 | 0.44 | 0.29 | 0.31 | |
| Estimated mean abundance (across survey sites) | 67.27 ± 27.15 (34.68, 152.85) | 24.19 ± 78.86 (1.50, 265.14) | 1.83 ± 2.25 (0.77, 6.94) | 0.62 ± 0.24 (0.35, 1.24) | 5.43 ± 10.93 (0.53, 32.18) | |
| Probability of individual detection (across survey sites) | ||||||
| Day | 0.007 ± 0.008 (0.002, 0.03) | 0.001 ± 0.007 (0.00, 0.001) | 0.22 ± 0.15 (0.03, 0.60) | 0.15 ± 0.13 (0.05, 0.53) | 0.02 ± 0.04 (0.00, 0.15) | |
| Night | 0.08 ± 0.05 (0.02, 0.21) | 0.11 ± 0.11 (0.005, 0.42) | 0.009 ± 0.02 (0.001, 0.06) | 0.02 ± 0.04 (0.00, 0.53) | 0.03 ± 0.04 (0.003, 0.12) | |
| Probability of individual detection: logit model | ||||||
| Intercept (day) | -5.31 ± 0.50 (-6.46, -4.50) | -8.61 ± 6.12 (-25.47, -4.83) | -1.30 ± 0.88 (-3.30, 0.16) | -1.67 ± 0.65 (-2.69, -0.40) | -4.01 ± 1.51 (-7.94, 1.92) | |
| Intercept (night) | -2.06 ± 0.5 (-3.25, -1.18) | -2.21 ± 1.26 (-5.35, -0.43) | -4.70 ± 1.22 (-7.69, -2.89) | -1.42 ± 0.59 (-2.69, 0.40) | -4.32 ± 1.51 (-8.19, -2.19) | |
| Ground temperature (standardised) | Degree Celsius 26.75 ± 3.61 (20.0-48.0) | 0.10 ± 0.10 (-0.11, 0.29) | -0.11 ± 0.56 (-1.64, 0.70) | -0.15 ± 0.19 (-0.55, 0.19) | -0.08 ± 0.34 (-0.92, 0.47) | 0.12 ± 0.34 (-0.56, 0.79) |
| Temperature 10cm above ground (standardised) | Degree Celsius 26.40 ± 2.47 (21.0-33.5) | 0.10 ± 0.14 (-0.19, 0.37) | -0.09 ± 0.16 (-1.29, 0.61) | -0.29 ± 0.37 (-1.18, 0.22) | 0.05 ± 0.40 (-0.75, 0.91) | 0.02 ± 0.44 (-0.88, 0.96) |
| Temperature 10cm underground (standardised) | Degree Celsius 26.40 ± 2.41 (21.6-33.8) | -0.11 ± 0.09 (-0.29, 0.06) | 0.12 ± 0.28 (-0.34, 0.81) | 0.07 ± 0.33 (-0.64, 0.71) | 0.35 ± 0.42 (-0.26, 1.37) | 0.43 ± 0.41 (-0.18, 1.39) |
| Temperature 120cm above ground (standardised) | Degree Celsius 26.25 ± 2.29 (21.40-33.80) | 0.52 ± 0.16 (0.22, 0.84) | 0.13 ± 0.47 (-0.54, 1.39) | 0.50 ± 0.52 (-0.25, 1.73) | 0.02 ± 0.44 (-0.96. 0.89) | -0.26 ± 0.54 (-1.62, 0.49) |
| Abundance: Negative Binomial regression | ||||||
| Distance to port (standardised) | Metres 1937.0 ± 2876.45 (138.0, 8575.0) | 0.01 ± 0.21 (-0.39, 0.43) | 0.26 ± 0.32 (-0.36, 0.98) | 0.15 ± 0.32 (-0.48, 0.79) | 0.01 ± 0.15 (-0.29, 0.35) | -0.04 ± 0.27 (-0.61, 0.44) |
| Cover of trees | Scale 2.00 ± 1.82 (0-5.00) | 0.12 ± 0.15 (-0.17, 0.43) | 0.07 ± 0.27 (-0.40, 0.70) | -0.25 ± 0.26 (-0.78, 0.23) | 0.01 ± 0.12 (-0.21, 0.27) | -0.27 ± 0.26 (-0.91, 0.10) |
| Cover of bush | Scale 2.00 ± 1.63 (0.00-5.00) | -0.11 ± 0.18 (-0.47, 0.47) | -0.13 ± 0.30 (-0.79, 0.62) | -0.32 ± 0.27 (-0.88, 0.19) | -0.05 ± 0.14 (-0.40, 0.18) | -0.01 ± 0.21 (-0.43, 0.44) |
| Cover of grass | Scale 4.00 ± 1.52 (0.00-5.00) | 0.48 ± 0.12 (0.24, 0.73) | 0.17 ± 0.25 (-0.28, 0.74) | 0.06 ± 0.19 (-0.33, 0.43) | -0.05 ± 0.11 (-0.28, 0.18) | 0.22 ± 0.26 (-0.12, 0.89) |
| Cover of rocks and stones | Scale 2.00 ± 0.97 (0.00-4.00) | -0.01 ± 0.24 (-0.48, 0.47) | -0.05 ± 0.35 (-0.80, 0.62) | 0.27 ± 0.34 (-0.32, 1.03) | -0.01 ± 0.15 (-0.33, 0.32) | 0.03 ± 0.27 (-0.48, 0.65) |
| Cover of human structures | Scale 3.50 ± 1.74 (0.00-5.00) | 0.69 ± 0.13 (0.46, 0.97) | 0.31 ± 0.24 (-0.07, 0.88) | 0.05 ± 0.21 (-0.31, 0.51) | -0.07 ± 0.12 (-0.35, 0.12) | 0.10 ± 0.19 (-0.26, 0.54) |
| Transect length (standardised) | Metres 175.00 ± 72.59 (60.00-384.00) | -0.01 ± 0.20 (-0.40, 0.41) | -0.01 ± 0.31 (-0.64, 0.62) | 0.33 ± 0.37 (-0.24, 1.18) | 0.01 ± 0.16 (-0.31, 0.35) | 0.14 ± 0.26 (-0.28, 0.74) |
| Elevation (standardised) | Metres above sea level 194.5 ± 94.25 (10.00, 303.00) | -0.05 ± 0.21 (-0.47, 0.36) | -0.01 ± 0.32 (-0.65, 0.64) | -0.69 ± 0.38 (-1.47. -0.01) | -0.05 ± 0.17 (-0.48, 0.21) | 0.01 ± 0.27 (-0.57. 0.55) |
| Overdispersion parameter Θ | 0.97 ± 0.30 (0.54, 1.70) | 0.31 ± 0.25 (0.12, 0.98) | 3.78 ± 15.17 (0.19,47.20) | 23.69 ± 14.70 (1.11, 48.68) | 24.97 ± 14.32 (1.66, 48.75) | |
Standardised Rapid Biological Assessment: a time-limited approach
We surveyed for alien reptiles during the dry season, 8th July to 7th August 2015, to minimise disturbances caused by the inclement weather conditions of the wet season. Each of the survey sites was surveyed using the time-limited transect approach; a standardised RBA for reptiles and amphibians (
Summary of the counts (number of observations per each 10-minute survey) of five species of alien reptiles on Christmas Island across the 34 survey sites (mean ± standard deviation, and range). Three repeated surveys were conducted during each day and night time conditions in each survey site. The raw data are available at https://figshare.com/s/e85ac13693bc6272437f
| Species | Day surveys | Night surveys |
| Common house gecko (Hemidactylus frenatus) | 0.74 ± 1.44 (0.00–7.00) | 6.51 ± 7.84 (0.00–35.00) |
| Stump-toed gecko (Gehyra mutilata) | 0.00 ± 0.00 (0.00–0.00) | 0.58 ± 1.45 (0.00–7.00) |
| Grass skink (Lygosoma bowringii) | 0.31 ± 0.84 (0.00–5.00) | 0.01 ± 0.10 (0.00–1.00) |
| Flowerpot snake (Indotyphlops braminus) | 0.08 ± 0.44 (0.00–2.00) | 0.11 ± 0.31 (0.00–1.00) |
| Wolf snake (Lycodon capucinus) | 0.06 ± 0.28 (0.00–2.00) | 0.05 ± 0.26 (0.00–2.00) |
Time-limited transects do not neccesarily require repeated survey occasions (
During each survey occasion, we recorded the number of individuals of each alien reptile species detected, and the temperature (ºC) at four different heights in the habitat (Table
Ndi,z ~ Binominal(Ni,pi,z) (1)
where Ni is the abundance of the species, and pi,z is the probability of individual detection (
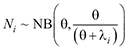 (2)
(2)
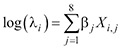 (3)
(3)
where βj (j = 1, …, 8) are the slopes of the model of the mean abundance, λi, in survey site i and Xi is a vector of the eight covariates used for modelling the mean abundance (Table
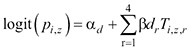 (4)
(4)
where αd are the time-of-day specific intercepts (day and night), βdr (r = 1, …, 4) are the slopes of the model of the probability of individual detection, pi,z, during survey occasion z at survey site i, and Ti,z is a vector of the four temperature covariates used for modelling the probability of individual detection (see Table
We employed Bayesian regularisation to construct robust model structures for the abundance and the probability of individual detection as a function of the covariates (
Results
Probabilities of individual detection of alien reptiles on Christmas Island
Our Bayesian regularised models were an adequate fit to the count data (Bayesian p-values close to 0.5 and non-skewed Q-Q plots in all cases; see Supplementary Methods), and revealed that the survey time (day vs night) was the main driver of the probability of individual detection across the five alien reptiles (Table

Estimated surveying effort required to detect all 10 individuals of each species of five alien reptiles in two situations and under a best-case detection scenario (i.e., sites surveyed during the time of the day when detection probabilities are higher). Top: surveying effort (minutes) to detect 10 individuals known to be present in one surveying site when only those 10 individuals are present; bottom: surveying effort (hours) to detect 10 individuals distributed at random across 34 surveying sites, where each occupied site harbours one individual, and only ten individuals are present across the 34 surveying sites. We used 10 individuals as an example to showcase and compare detection efforts across different situations. Best-case detection scenarios are night time surveys (common house gecko, stump-toed gecko, and flowerpot snake) and daytime surveys (grass skink and wolf snake). Each dot represents a realisation from 1000 simulations.
All the posterior estimates of the probabilities of individual detection were relatively low, with the upper 95% Credible Interval estimates of daytime surveys of the grass skink and night time surveys of flowerpot snakes being the only values exceeding 0.5 (Table
Abundance of alien reptiles on Christmas Island
Common house geckos were the most frequently encountered species, followed by stump-toed geckos during night time surveys and grass skinks during daytime surveys (Table
Our Negative Binomial regressions revealed that common house geckos were more common in sites with higher grass and human structure cover (positive relationships), and that grass skinks tended to become rarer in higher elevation sites (negative relationship with elevation). Across the five alien reptiles, all the covariates had uncertain effects on the estimated alien reptile abundances and probabilities of detection, with wide posterior estimates overlapping zero (Table
Discussion
The probabilities of detecting individuals using a standardised RBA were consistently low across all the alien reptiles found on Christmas Island. Both the detectability and the abundance of those alien reptiles were difficult to explain given the rather uncertain effects of the covariates tested in our models, even when our surveys explored a representative and variable sample of environmental conditions (Table
It is particularly important to protect island ecosystems, where alien reptiles have produced substantial negative impacts and whose native biodiversity is highly exposed to the threat of alien species (
The compounded effects of low detection probabilities and uncertain effects of covariates will hinder effective measures to manage the emergence of alien reptiles, a conclusion reinforced by previous species-specific research into the management of invasive brown snakes (Boiga irregularis) in Guam (
Framed in this context of overarching uncertainties and high costs, strong preventive policies should be a priority to address the emergent threat of alien reptiles on islands and elsewhere (
Preventive management activities should be complemented with early detection surveys aimed at detecting new populations of alien reptiles promptly (
More broadly, biosecurity regulations, strict quarantine, and early detection activities should be considered within the framework of robust anticipatory policy-making (
The implementation of stringent preventive policies and early detection activities might be more straightforward on oceanic islands, where their remoteness commonly requires all goods and commodities to be imported via shipping and air traffic routes arriving in a small number (usually one) of ports and airports, limiting the number of potential pathways of transport and points of entry into the island (
We have focussed on alien reptiles on islands in this research, but our conclusions are likely applicable to other groups of emergent alien species globally; such as vertebrates in the pet trade (
Acknowledgements
This work would not have been possible without the support provided by the all the staff of the Christmas Island National Park, the Christmas Island Natural Resource Management Board, and the Department of Agriculture and Water Resources (Indian Ocean). This research was conducted under permit number CIN_2015_5 (Christmas Island National Park, Director of National Parks, Australian Government) and a Licence to undertake research activities on regulated crown land. The research was approved by the University of Adelaide Animal Ethics Committee (S-2014-155). Andrew Robinson, John Measey, and an anonymous reviewer provided constructive feedback that helped improve this manuscript. PG-D was supported by an IPRS/APA scholarship (DET), an Invasive Animals CRC PhD scholarship, and the IA CRC Student Grant. This work was supported by an ARC Discovery Grant (DP140102319) to JVR and PC and ARC Future Fellowships to PC and JVR (FT0991420, FT130100254).
References
- Anderson DP, Ramsey DSL, Nugent G, Bosson M, Livingstone P, Martin PAJ, Sergeant E, Gormley AM, Warburton B (2013) A novel approach to assess the probability of disease eradication from a wild-animal reservoir host. Epidemiology & Infection 141: 1509–1521. https://doi.org/10.1017/S095026881200310X
- Barker RJ, Schofield MR, Link WA, Sauer JR (2018) On the reliability of N-mixture models for count data. Biometrics 74: 369–377. https://doi.org/10.1111/biom.12734
- Bellard C, Rysman J-F, Leroy B, Claud C, Mace GM (2017) A global picture of biological invasion threat on islands. Nature Ecology & Evolution 1: 1862–1869. https://doi.org/10.1038/s41559-017-0365-6
- Boston J (2016) Anticipatory governance: how well is New Zealand safeguarding the future? Policy Quarterly 12: 11–24. https://doi.org/10.26686/pq.v12i3.4614
- Boston J (2017) Safeguarding the Future: Governing in an Uncertain World. Bridget Williams Books, Wellington, 215 pp. https://doi.org/10.7810/9780947518257
- Brenton-Rule E, Frankel S, Lester P (2016) Improving management of invasive species: New Zealand’s approach to pre-and post-border pests. Policy Quarterly 12: 17–25. https://doi.org/10.26686/pq.v12i1.4582
- Capinha C, Seebens H, Cassey P, García-Díaz P, Lenzner B, Mang T, Moser D, Pyšek P, Rödder D, Scalera R, Winter M, Dullinger S, Essl F (2017) Diversity, biogeography and the global flows of alien amphibians and reptiles. Diversity and Distributions 23: 1313–1322. https://doi.org/10.1111/ddi.12617
- Catenazzi A, Richards S, Glos J (2016) Herpetofauna. In: Larsen TH (Ed.) Core Standardized Methods for Rapid Biological Field Assessment. Conservation International, Arlington, VA, 110–126.
- Chapple DG, Knegtmans J, Kikillus H, van Winkel D (2016) Biosecurity of exotic reptiles and amphibians in New Zealand: building upon Tony Whitaker’s legacy. Journal of the Royal Society of New Zealand 46: 66–84. https://doi.org/10.1080/03036758.2015.1108344
- Christy MT, Yackel Adams AA, Rodda GH, Savidge JA, Tyrrell CL (2010) Modelling detection probabilities to evaluate management and control tools for an invasive species. Journal of Applied Ecology 47: 106–113. https://doi.org/10.1111/j.1365-2664.2009.01753.x
- Claussen J (2005) Native Plants of Christmas Island. Australian Biological Resources Study, CSIRO, Canberra, Australia, 152 pp.
- Conn P, Johnson D, Williams P, Melin S, Hooten M (2018) A guide to Bayesian model checking for ecologists. Ecological Applications 88: 526–542. https://doi.org/10.1002/ecm.1314
- Conroy MJ, Peterson JT (2013) Decision Making in Natural Resource Management: a Structured, Adaptive Approach. John Wiley & Sons, West Sussex, 474 pp. https://doi.org/10.1002/9781118506196
- de Valpine P, Turek D, Paciorek CJ, Anderson-Bergman C, Lang DT, Bodik R (2017) Programming with models: writing statistical algorithms for general model structures with NIMBLE. Journal of Computational and Graphical Statistics 26: 403–413. https://doi.org/10.1080/10618600.2016.1172487
- Dorcas ME, Willson JD, Reed RN, Snow RW, Rochford MR, Miller MA, Meshaka WE, Andreadis PT, Mazzotti FJ, Romagosa CM (2012) Severe mammal declines coincide with proliferation of invasive Burmese pythons in Everglades National Park. Proceedings of the National Academy of Sciences 109: 2418–2422. https://doi.org/10.1073/pnas.1115226109
- Dunn PK, Smyth GK (1996) Randomized quantile residuals. Journal of Computational and Graphical Statistics 5: 236–244. https://doi.org/10.2307/1390802
- Durso AM, Seigel RA (2015) A snake in the hand is worth 10,000 in the bush. Journal of Herpetology 49: 503–506. https://doi.org/10.1670/15-49.04.1
- Durso AM, Willson JD, Winne CT (2011) Needles in haystacks: Estimating detection probability and occupancy of rare and cryptic snakes. Biological Conservation 144: 1508–1515. https://doi.org/10.1016/j.biocon.2011.01.020
- Early R, Bradley BA, Dukes JS, Lawler JJ, Olden JD, Blumenthal DM, Gonzalez P, Grosholz ED, Ibanez I, Miller LP, Sorte CJB, Tatem AJ (2016) Global threats from invasive alien species in the twenty-first century and national response capacities. Nature Communications 7: 12485. https://doi.org/10.1038/ncomms12485
- Ficetola GF, Barzaghi B, Melotto A, Muraro M, Lunghi E, Canedoli C, Parrino EL, Nanni V, Silva-Rocha I, Urso A (2018) N-mixture models reliably estimate the abundance of small vertebrates. Scientific Reports 8: 10357. https://doi.org/10.1038/s41598-018-28432-8
- Fritts TH (1993) The common wolf snake, Lycodon aulicus capucinus, a recent colonists of Christmas Island in the Indian Ocean. Wildlife Research 20: 261–265. https://doi.org/10.1071/WR9930261
- Gallo T, Waitt D (2011) Creating a successful citizen science model to detect and report invasive species. BioScience 61: 459–465. https://doi.org/10.1525/bio.2011.61.6.8
- García-Díaz P, Ramsey DSL, Woolnough AP, Franch M, Llorente GA, Montori A, Buenetxea X, Larrinaga AR, Lasceve M, Álvarez A, Traverso JM, Valdeón A, Crespo A, Rada V, Ayllón E, Sancho V, Lacomba JI, Bataller JV, Lizana M (2017a) Challenges in confirming eradication success of invasive red-eared sliders. Biological Invasions 19: 2739–2750. https://doi.org/10.1007/s10530-017-1480-7
- García-Díaz P, Ross JV, Woolnough AP, Cassey P (2017b) Managing the risk of wildlife disease introduction: pathway-level biosecurity for preventing the introduction of alien ranaviruses. Journal of Applied Ecology 54: 234–241. https://doi.org/10.1111/1365-2664.12749
- Gregory S, Henderson W, Smee E, Cassey P (2014) Eradications of Vertebrate Pests in Australia: a Review and Guidelines for Future Best Practice. Invasive Animals Cooperative Research Centre, Canberra, 76 pp.
- Guillera-Arroita G, Lahoz-Monfort JJ, van Rooyen AR, Weeks AR, Tingley R (2017) Dealing with false positive and false negative errors about species occurrence at multiple levels. Methods in Ecology and Evolution 8: 1081–1091. https://doi.org/10.1111/2041-210X.12743
- Hartig F (2018) Package “DHARMa”: residual diagnostics for hierarchical (multi-level/mixed) regression models. Package “DHARMa” https://cran.r-project.org/web/packages/DHARMa/vignettes/DHARMa.html
- Helmus MR, Mahler DL, Losos JB (2014) Island biogeography of the Anthropocene. Nature 513: 543–546. https://doi.org/10.1038/nature13739
- Hobbs NT, Hooten MB (2015) Bayesian Models: a Statistical Primer for Ecologists. Princeton University Press, Princeton, 299 pp. https://doi.org/10.1515/9781400866557
- Holden MH, Nyrop JP, Ellner SP (2016) The economic benefit of time-varying surveillance effort for invasive species management. Journal of Applied Ecology 53: 712–721 https://doi.org/10.1111/1365-2664.12617
- Hooten MB, Hobbs NT (2015) A guide to Bayesian model selection for ecologists. Ecological Monographs 85: 3–28. https://doi.org/10.1890/14-0661
- Howald G, Donlan C, Galván JP, Russell JC, Parkes J, Samaniego A, Wang Y, Veitch D, Genovesi P, Pascal M (2007) Invasive rodent eradication on islands. Conservation Biology 21: 1258–1268. https://doi.org/10.1111/j.1523-1739.2007.00755.x
- Hulme PE (2015) Invasion pathways at a crossroad: policy and research challenges for managing alien species introductions. Journal of Applied Ecology 52: 1418–1424. https://doi.org/10.1111/1365-2664.12470
- Hulme PE, Bacher S, Kenis M, Klotz S, Kühn I, Minchin D, Nentwig W, Olenin S, Panov V, Pergl J, Pyšek P, Roques A, Sol D, Solarz W, Vilá M (2008) Grasping at the routes of biological invasions: a framework for integrating pathways into policy. Journal of Applied Ecology 45: 403–414. https://doi.org/10.1111/j.1365-2664.2007.01442.x
- Hunter ME, Oyler-McCance SJ, Dorazio RM, Fike JA, Smith BJ, Hunter CT, Reed RN, Hart KM (2015) Environmental DNA (eDNA) sampling improves occurrence and detection estimates of invasive Burmese pythons. PloS ONE 10: e0121655. https://doi.org/10.1371/journal.pone.0121655
- Jarrad FC, Barrett S, Murray J, Stoklosa R, Whittle P, Mengersen K (2011) Ecological aspects of biosecurity surveillance design for the detection of multiple invasive animal species. Biological Invasions 13: 803–818. https://doi.org/10.1007/s10530-010-9870-0
- Jarrad FC, Low-Choy S, Mengersen K (2015) Biosecurity Surveillance: Quantitative Approaches. CABI, Croydon, 386 pp. https://doi.org/10.1079/9781780643595.0000
- Kéry M (2002) Inferring the absence of a species: a case study of snakes. Journal of Wildlife Management 66: 330–338. https://doi.org/10.2307/3803165
- Kéry M (2018) Identifiability in N-mixture models: a large-scale screening test with bird data. Ecology 99: 281–288. https://doi.org/10.1002/ecy.2093
- Kéry M, Royle AJ (2016) Applied Hierarchical Modeling in Ecology. Analysis of Distribution, Abundance and Species Richness in R and BUGS. Academic Press, London, 783 pp. https://doi.org/10.1016/B978-0-12-801378-6.00001-1
- Kraus F (2009) Alien Reptiles and Amphibians: A Scientific Compendium and Analysis. Invading Nature: Springer Series in Invasion Ecology. Springer, Dordrecht, 369 pp.
- Kraus F (2015) Impacts from invasive reptiles and amphibians. Annual Review of Ecology, Evolution, and Systematics 46: 75–97. https://doi.org/10.1146/annurev-ecolsys-112414-054450
- Larsen TH (2016) Core Standardized Methods for Rapid Biological Field Assessment. Conservation International, Arlington, 208 pp.
- Lawson B, Petrovan SO, Cunningham AA (2015) Citizen science and wildlife disease surveillance. EcoHealth 12: 693–702. https://doi.org/10.1007/s10393-015-1054-z
- Leung B, Springborn MR, Turner JA, Brockerhoff EG (2014) Pathway-level risk analysis: the net present value of an invasive species policy in the US. Frontiers in Ecology and the Environment 12: 273–279. https://doi.org/10.1890/130311
- Lodge DM, Simonin PW, Burgiel SW, Keller RP, Bossenbroek JM, Jerde CL, Kramer AM, Rutherford ES, Barnes MA, Wittmann ME, Chadderton WL, Apriesnig JL, Beletsky D, Cooke RM, Drake JM, Egan SP, Finnoff DC, Gantz CA, Grey EK, Hoff MH, Howeth JG, Jensen RA, Larson ER, Mandrak NE, Mason DM, Martinez FA, Newcomb TJ, Rothlisberger JD, Tucker AJ, Warziniack TW, Zhang H (2016) Risk analysis and bioeconomics of invasive species to inform policy and management. Annual Review of Environment and Resources 41: 453–488. https://doi.org/10.1146/annurev-environ-110615-085532
- McCarthy MA, Moore JL, Morris WK, Parris KM, Garrard GE, Vesk PA, Rumpff L, Giljohann KM, Camac JS, Bau SS, Friend T, Harrison B, Yue B (2013) The influence of abundance on detectability. Oikos 122: 717–726. https://doi.org/10.1111/j.1600-0706.2012.20781.x
- McIntosh CR, Finnoff DC, Settle C, Shogren JF (2009) Economic valuation and invasive species. In: Keller RP, Lodge DM, Lewis MA, Shogren JF (Eds) Bioeconomics Invasive Species: Integrating Ecology, Economics, Policy, and Management. Oxford University Press, New York, 151–179.
- Meyerson LA, Reaser JK (2002) Biosecurity: moving toward a comprehensive approach. BioScience 52: 593–600.
- Morris TP, White IR, Crowther JR (2017) Using simulation studies to evaluate statistical methods. ArXiv Prepr. ArXiv171203198.
- Moser D, Lenzner B, Weigelt P, Dawson W, Kreft H, Pergl J, Pyšek P, van Kleunen M, Winter M, Capinha C, Cassey P, Dullinger S, Economo EP, García-Díaz P, Guénard B, Hofhansl F, Mang T, Seebens H, Essl F (2018) Remoteness promotes biological invasions on islands worldwide. Proceedings of the National Academy of Sciences 115: 9270–9275. https://doi.org/10.1073/pnas.1804179115
- Powell R (2011) Conservation of the herpetofauna on the Dutch Windward Islands: St. Eustatius, Saba, and St. Maarten. In: Hailey A, Wilson BS, Horrocks JA (Eds) Conservation of Caribbean Island Herpetofaunas, Vol. 2: Regional Accounts of the West Indies. Brill, Leiden, The Netherlands, 189–204. https://doi.org/10.1163/ej.9789004194083.i-439.76
- R Development Core Team (2015) R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria.
- Ramsey DS, Parkes J, Morrison SA (2009) Quantifying eradication success: the removal of feral pigs from Santa Cruz Island, California. Conservation Biology 23: 449–459. https://doi.org/10.1111/j.1523-1739.2008.01119.x
- Reed RN, Hart KM, Rodda GH, Mazzotti FJ, Snow RW, Cherkiss M, Rozar R, Goetz S (2011) A field test of attractant traps for invasive Burmese pythons (Python molurus bivittatus) in southern Florida. Wildlife Research 38: 114–121. https://doi.org/10.1071/WR10202
- Rodda GH, Dean-Bradley K, Campbell EW, Fritts TH, Lardner B, Adams AAY, Reed RN (2015) Stability of detectability over 17 years at a single site and other lizard detection comparisons from Guam. Journal of Herpetology 49: 513–521. https://doi.org/10.1670/14-085
- Rodda GH, Sawai Y, Chiszar D, Tanaka H (1999) Problem Snake Management: The Habu and the Brown Treesnake. Comstock Publishing Associates, Ithaca, 534 pp.
- Rout TM, Moore JL, McCarthy MA (2014) Prevent, search or destroy? A partially observable model for invasive species management. Journal of Applied Ecology 51: 804–813. https://doi.org/10.1111/1365-2664.12234
- Royle JA (2004) N‐mixture models for estimating population size from spatially replicated counts. Biometrics 60: 108–115. https://doi.org/10.1111/j.0006-341X.2004.00142.x
- Scott JK, McKirdy SJ, van der Merwe J, Green R, Burbidge AA, Pickles G, Hardie DC, Morris K, Kendrick PG, Thomas ML, Horton KL, O’Connor SM, Downs J, Stoklosa R, Lagdon R, Marks B, Nairn M, Mengersen K (2017) Zero-tolerance biosecurity protects high-conservation-value island nature reserve. Scientific Reports 7: 772. https://doi.org/10.1038/s41598-017-00450-y
- Seebens H, Blackburn TM, Dyer EE, Genovesi P, Hulme PE, Jeschke JM, Pagad S, Pyšek P, van Kleunen M, Winter M, Ansong M, Arianoutsou M, Bacher S, Blasius B, Brockerhoff EG, Brundu G, Capinha C, Causton CE, Celesti-Grapow L, Dawson W, Dullinger S, Economo EP, Fuentes N, Guénard B, Jäger H, Kartesz J, Kenis M, Kühn I, Lenzner B, Liebhold AM, Mosena A, Moser D, Nentwig W, Nishino M, Pearman D, Pergl J, Rabitsch W, Rojas-Sandoval J, Roques A, Rorke S, Rossinelli S, Roy HE, Scalera R, Schindler S, Štajerová K, Tokarska-Guzik B, Walker K, Ward DF, Yamanaka T, Essl F (2018) Global rise in emerging alien species results from increased accessibility of new source pools. Proceedings of the National Academy of Sciences 115: 2264–2273. https://doi.org/10.1073/pnas.1719429115
- Sewell D, Guillera-Arroita G, Griffiths RA, Beebee TJC (2012) When is a species declining? Optimizing survey effort to detect population changes in reptiles. PloS ONE 7: e43387. https://doi.org/10.1371/journal.pone.0043387
- Silva-Rocha I, Salvi D, Sillero N, Mateo JA, Carretero MA (2015) Snakes on the Balearic Islands: an invasion tale with implications for native biodiversity conservation. PloS ONE 10: e0121026. https://doi.org/10.1371/journal.pone.0121026
- Smith MJ, Boland CR, Maple D, Tiernan B (2012a) The Christmas Island blue-tailed skink (Cryptoblepharus egeriae): a survey protocol and an assessment of factors that relate to occupancy and detection. Records of the Western Australia Museum 427: 40–44. https://doi.org/10.18195/issn.0312-3162.27(1).2012.040-044
- Smith MJ, Cogger H, Tiernan B, Maple D, Boland C, Napier F, Detto T, Smith P (2012b) An oceanic island reptile community under threat: The decline of reptiles on Christmas Island, Indian Ocean. Herpetological Conservation and Biology 7: 206–218.
- Sutherland WJ (2006) Ecological Census Techniques: A Handbook. Cambridge University Press, Cambridge, 446 pp. https://doi.org/10.1017/CBO9780511790508
- Vander-Zanden MJ, Hansen GJ, Higgins SN, Kornis MS (2010) A pound of prevention, plus a pound of cure: early detection and eradication of invasive species in the Laurentian Great Lakes. Journal of Great Lakes Research 36: 199–205. https://doi.org/10.1016/j.jglr.2009.11.002
- van Wilgen NJ, Gillespie MS, Richardson DM, Measey J (2018) A taxonomically and geographically constrained information base limits non-native reptile and amphibian risk assessment: a systematic review. PeerJ 6: e5850. https://doi.org/10.7717/peerj.5850
- Wyatt KB, Campos PF, Gilbert MTP, Kolokotronis S-O, Hynes WH, DeSalle R, Daszak P, MacPhee RD, Greenwood AD (2008) Historical mammal extinction on Christmas Island (Indian Ocean) correlates with introduced infectious disease. PloS ONE 3: e3602. https://doi.org/10.1371/journal.pone.0003602
- Yemshanov D, Koch FH, Ducey M, Venette RC (2015) Making Invasion models useful for decision makers; incorporating uncertainty, knowledge gaps, and decision-making preferences. In: Venette RC (Ed.) Pest Risk Modeling and Mapping for Invasive Alien Species. CABI, Wallingford, 206–222. https://doi.org/10.1079/9781780643946.0206
Supplementary material
Model evaluation and estimability
Data type: measurement