Research Article |
|
Corresponding author: Ashlyn L. Padayachee ( ashlyn.levadia@gmail.com ) Academic editor: Ramiro Bustamante
© 2019 Ashlyn L. Padayachee, Şerban Procheş, John R. U. Wilson.
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Padayachee AL, Procheş Ş, Wilson JRU (2019) Prioritising potential incursions for contingency planning: pathways, species, and sites in Durban (eThekwini), South Africa as an example. NeoBiota 47: 1-21. https://doi.org/10.3897/neobiota.47.31959
|
Abstract
Increased trade and travel have resulted in an increasing rate of introduction of biological organisms to new regions. Urban environments, such as cities, are hubs for human activities facilitating the introduction of alien species. Additionally, cities are susceptible to invading organisms as a result of the highly altered and transformed nature of these environments. Despite best efforts at prevention, new incursions of alien species will occur; therefore, prioritising incursion response efforts is essential. This study explores these ideas to identify priorities for strategic prevention planning in a South African city, Durban (eThekwini), by combining data from alien species watch lists, environmental criteria, and the pathways which facilitate the introduction of alien species in the city. Three species (with known adverse impacts elsewhere in the world) were identified as highly likely to be introduced and established in Durban (Alternanthera philoxeroides, Lithobates catesbeianus and Solenopsis invicta). These species are most likely to enter at either the Durban Harbour; pet and aquarium stores; or plant nurseries and garden centres – therefore active surveillance should target these sites as well as adjacent major river systems and infrastructure. We suggest that the integrated approach (species, pathways, and sites) demonstrated in this study will help prioritise resources to detect the most likely and damaging future incursions of alien species.
Keywords
biological invasions, early detection, incursion response planning, prioritisation, alligator weed, southern sandbur, American bullfrog, red imported fire ant
Introduction
Human-related activities such as trade and travel have facilitated the increased introduction of biological organisms outside their native range (
Similarly, pathways facilitating the introduction of alien species to new regions need to be identified and the risk associated with introductions facilitated through these pathways assessed. Priority should then be given to the pathways of introduction which pose the highest risk of facilitating the introduction of alien species (
Finally, sites are assessed as high-risk based on the likelihood of an invasion (i.e., the exposure to incursions and whether incursions will establish themselves and become invasions) and sensitivity (i.e., most vulnerable to the impacts of invasions) (
In this study we identify potential future incursions in Durban (eThekwini), South Africa, based on selected alien species, the pathways facilitating their introduction, and the sites most at risk of being invaded by these species. By jointly considering species, pathways, and sites, we aim to provide a tool for decision makers to more effectively target surveillance and contingency planning.
Methods
The eThekwini municipality is one of the largest port cities on the east coast of the African continent and is an important economic centre in South Africa (
To develop a methodology for decision makers to assign priorities for prevention strategies we: 1) identified cities with a similar climate to eThekwini; 2) used existing lists of species considered as not present in South Africa that pose an unacceptable risk of invasion; 3) identified which of the selected species are likely to have pathways facilitating their introduction to eThekwini; 4) developed climatic suitability models for the selected species based on the climate in eThekwini; and 5) linked the climate and pathway information to identify sites within eThekwini that should be the focus of contingency planning for particular species (Figure

A simple and rapid method to prioritise targets for contingency planning to prevent biological invasions. The method identifies priority sites for managing particular high-risk incursions. A Shows the selection criteria used to select target species for climatic suitability analyses, with the number of species selected at each stage of selection indicated in parentheses. B Shows the criteria used to identify potential points of introduction for the select target species, as well as the criteria used to identify potential points of naturalisation, i.e. priority sites for monitoring in the eThekwini municipality.
Human population, as a result of the associated activities (trade and travel), is one of the main correlates of species introductions into regions outside of their native range (
The National Environmental Management: Biodiversity Act (No. 10 of 2004) governs all biodiversity related issues in South Africa, including biological invasions (NEMBA, 2014). In regulations under NEMBA, a prohibited species list was created, based in part on expert opinion, that lists species that are not believed to be present in South Africa and whose introduction should be prevented (DEA, 2016). The implication is that strategic prevention plans should be developed for all species on the prohibited list. Separate to this,
We used these national lists and applied our own selection criteria (Figure
Maximum entropy distribution modelling was selected to map the potential geographic distribution and evaluate the risk of invasion of the remaining species (Maxent v3.4.1 –
Results
Fifty-nine species were on both the NEMBA prohibited species list and the watch list produced by
We identified the pathways of introduction for each of the remaining species. At this stage, we excluded V. vulpes (red fox) as it is extremely unlikely to be introduced by the only pathways that have historically led to its introduction to other countries (hunting in the wild and fur farms – GISD, 2018). The pathways facilitating the introduction of C. echinatus were unknown (GISD, 2018). This meant that while it was possible to still build a climatic suitability model for the species, it is not possible, at this stage, to link climate suitability to introduction pathways (Box
Pathways of introduction, preferred habitats, potential entry points, sites to monitor, and climatic suitability for Alternanthera philoxeroides (alligator weed).
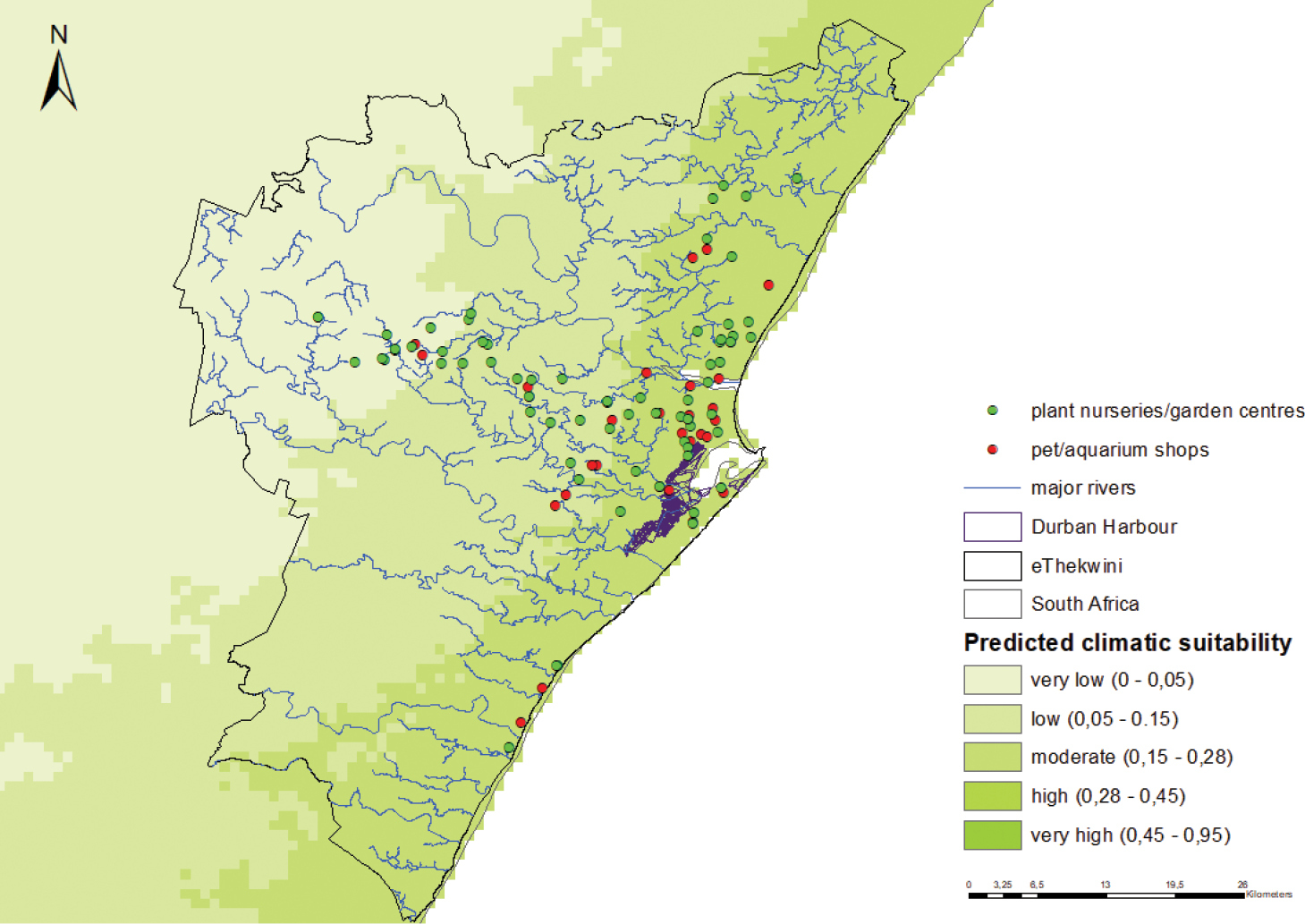
Figure B1. Predicted climatic suitability A. philoxeroides in Durban. The model is highly accurate in predicting climatic suitability (0.929 ± 0.007 – AUC±SD). Predicted suitability is indicated using a colour scale (darker shades indicate higher predicted suitability). Also indicated are the potential points of introduction and potential points of first naturalisation to monitor for A. philoxeroides in Durban. Pathways of introduction: Ship ballast (historical), transportation of habitat material, ornamental purposes Potential points of first introduction: The Durban harbour, plant nurseries and garden centres, pet and aquarium shops Habitat and Land uses: Alternanthera philoxeroides can grow in a variety of habitats but is usually found in aquatic habitats, particularly rivers, lakes, dams, ponds, canals, flood plains and irrigation channels Habitats present in Durban: Yes Potential sites of first naturalisation in Durban: The Durban harbour and adjacent river systems (particularly uMhlatuzana and uMbilo river systems) |
Species distribution models
The climate models developed for the selected species ranged from highly accurate model performance to moderately accurate performance based on the AUC of receiver operating characteristics (see Table
Pathways of introduction, preferred habitats, potential entry points, sites to monitor, and climatic suitability for Cenchrus echinatus (southern sandbur).
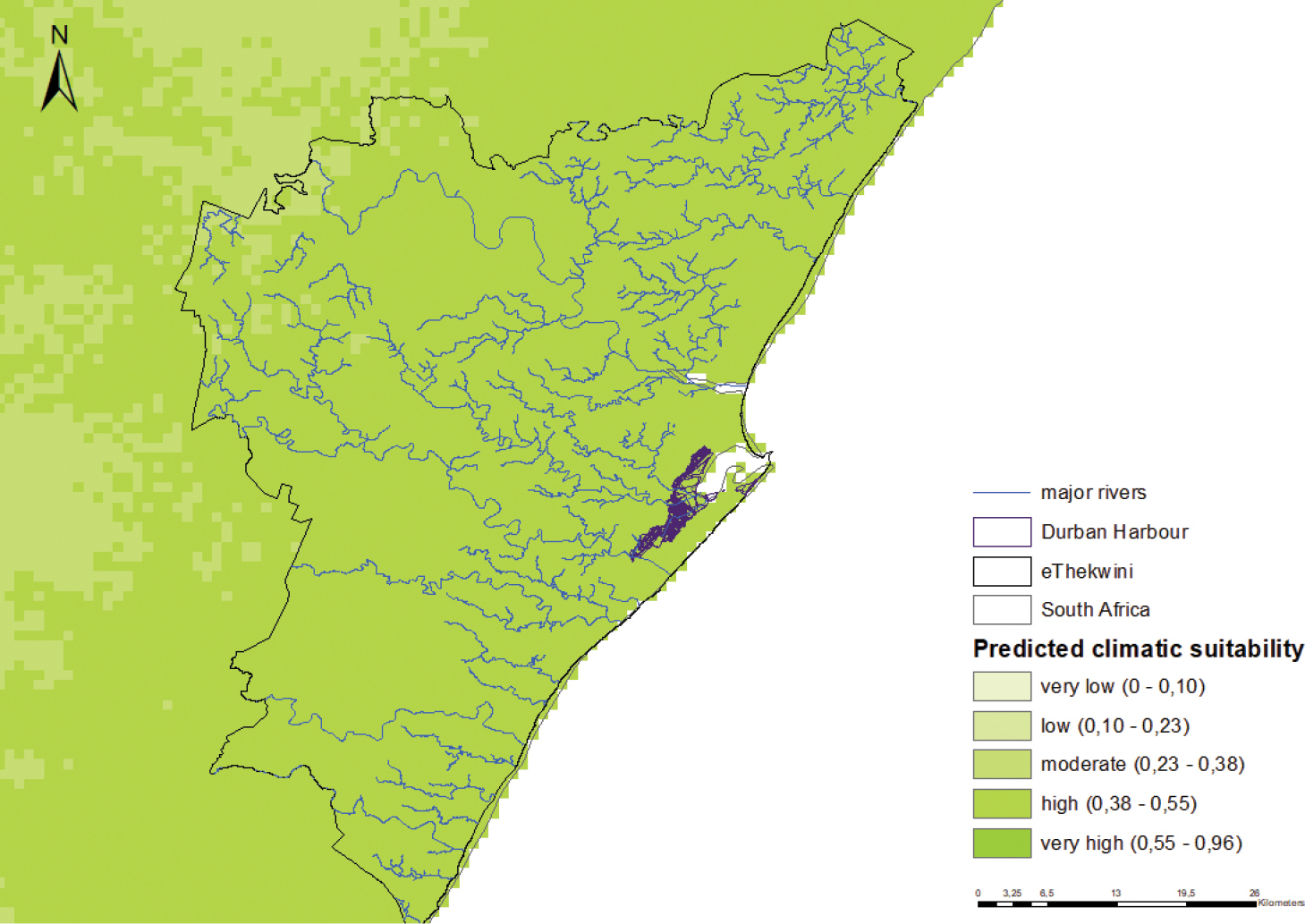
Figure B2. Predicted climatic suitability for C. echinatus in Durban. The model is moderately accurate in predicting climatic suitability (0.812 ± 0.008 – AUC±SD). Predicted climatic suitability is indicated using a colour scale (darker shades indicate higher predicted suitability). Even though pathways of introduction for this species could not be identified with certainty, the potential points of introduction and first naturalisation (i.e. where to monitor) for C. echinatus in Durban are indicated. Pathways of introduction: Unknown Potential points of first introduction: The Durban harbour Habitat and Land uses: Cenchrus echinatus favours temperate and tropical zones. This species is usually found in open lands, cultivated fields, along roadsides and coastal environments and waste places. Habitats present in Durban: Yes Potential sites of first naturalisation: The Durban harbour and adjacent beach environments and sand dunes |
Additionally, we superimposed pet and aquarium shops, nurseries and garden centres, the major river systems and the Durban Harbour data with the climatic suitability models (see Boxes
Pathways of introduction, preferred habitats, potential entry points, sites to monitor, and climatic suitability for Lithobates catesbeianus.
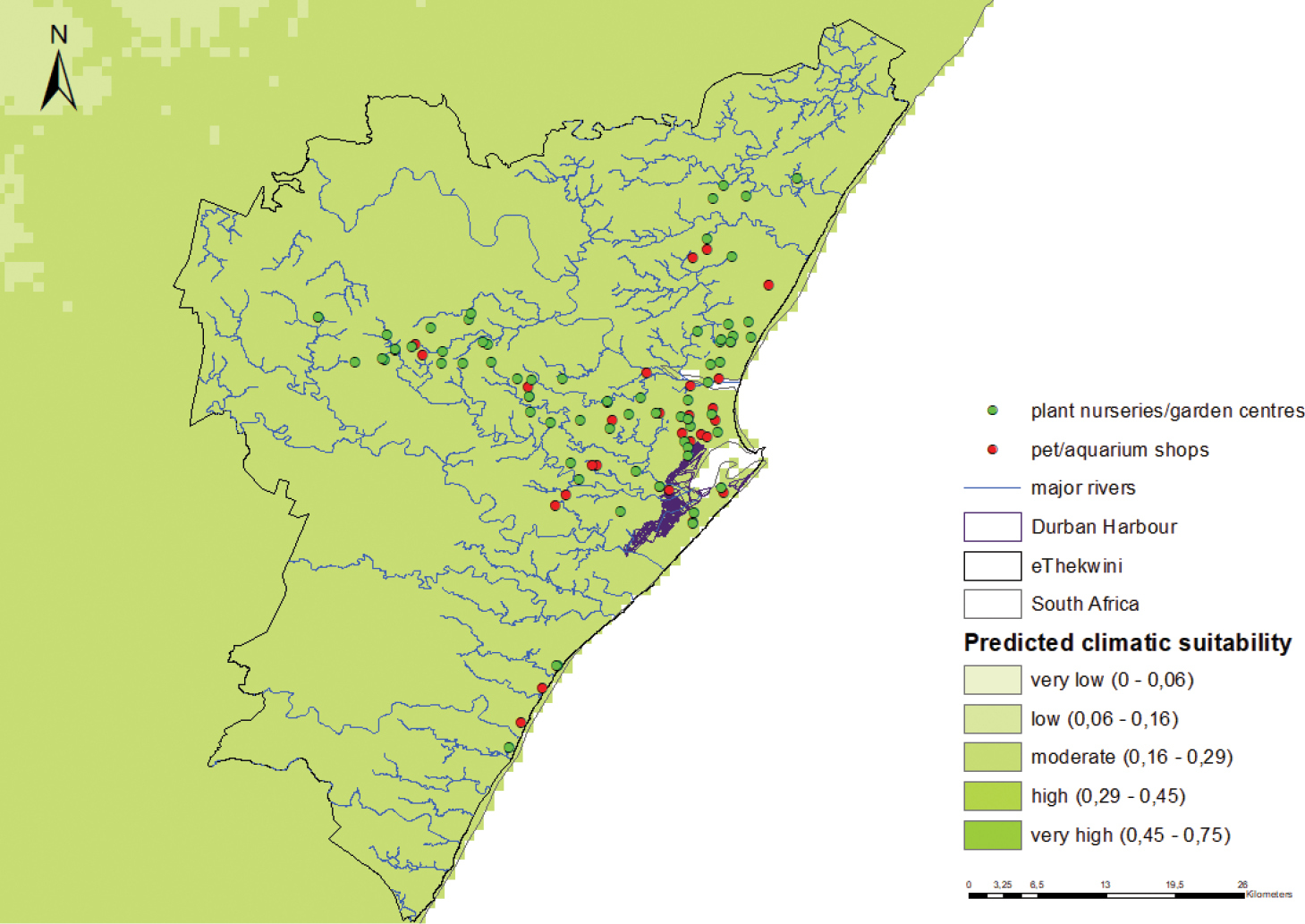
Figure B3: Predicted climatic suitability of L. catesbeianus in Durban. The model is moderately accurate in predicting climatic suitability (0.791 ± 0.005 – AUC±SD). Predicted suitability is indicated using a colour scale (darker shades indicate higher predicted suitability). Also indicated are the potential points of first naturalisation (i.e. priorities for monitoring) for C. echinatus in Durban. Pathways of introduction: Biological control, landscape; floral and faunal improvement, release in use for nature, aquaculture (food source), ornamental purposes Potential points of first introduction: The Durban harbour, pet and aquarium shops Habitat and Land uses: Lithobates catesbeianus prefers warm, moist environments and requires permanent, shallow and still bodies of water. This frog species usually occupies ponds, swamps, streams and irrigation ditches Habitats present in Durban: Yes Potential sites of first naturalisation: Major river systems, especially those adjacent to potential points of introduction (pet and aquarium shops) |
pathways of introduction, preferred habitats, potential entry points, sites to monitor, and climatic suitability for Solenopsis invicta.
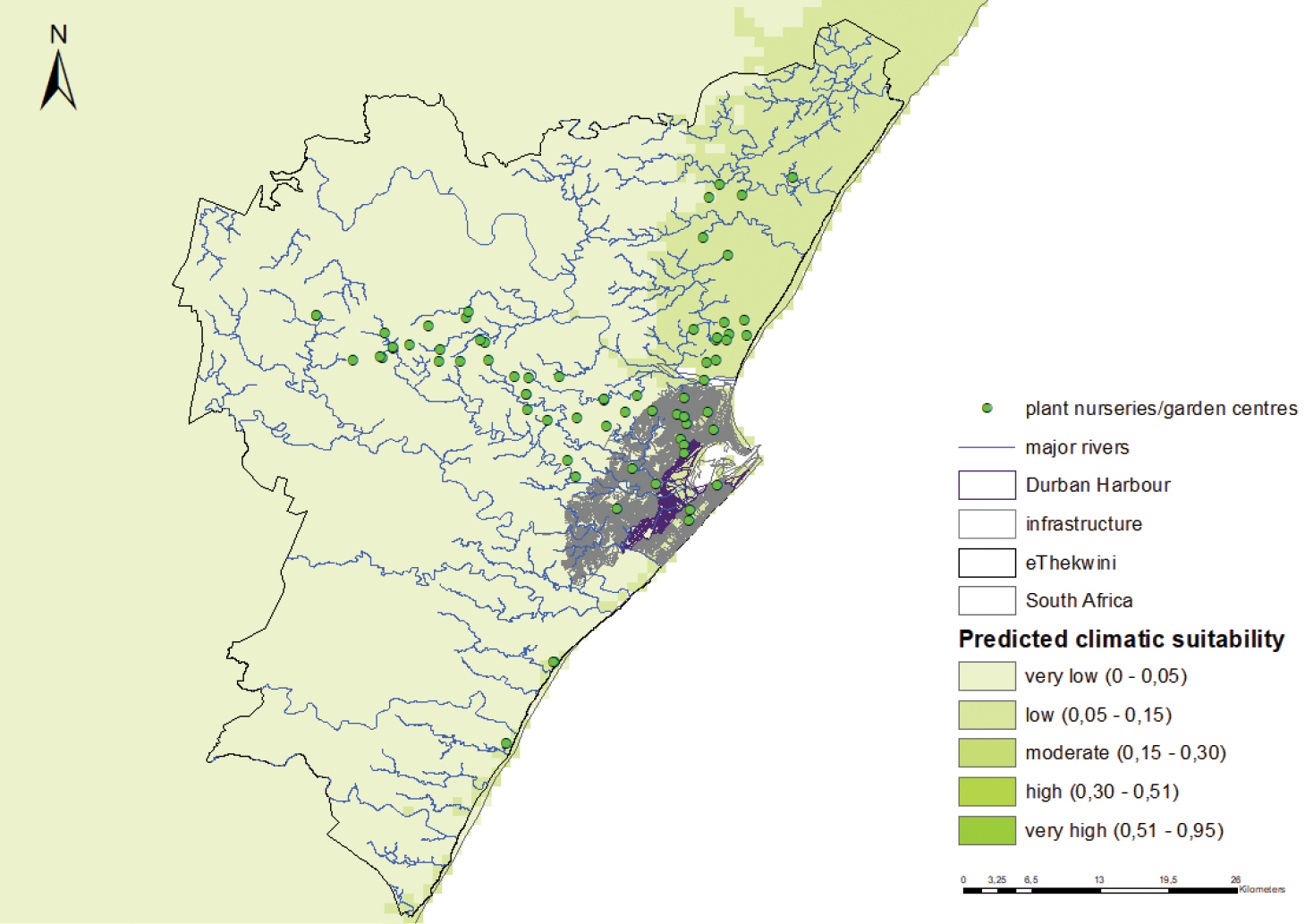
Figure B4: Predicted climatic suitability of S. invicta in Durban. The model is highly accurate in predicting climatic suitability (0.961 ± 0.006 – AUC±SD). Predicted suitability is indicated using a colour scale (darker shades indicated higher predicted suitability). Also indicated are the potential points of introduction and fist naturalisation to monitor for S. invicta in Durban. Pathways of introduction: Contaminated nursery material, translocation of machinery and equipment, organic wood packaging Potential points of first introduction: The Durban harbour, plant and nursery material Habitat and Land uses: Solenopsis invicta can occupy a wide variety of habitats and can become dominant in altered habitats. This ant species is found in disturbed or developed forests or on trails near buildings Habitats present in Durban: Yes Potential sites of first naturalisation: The Durban harbour and adjacent built infrastructure, plant nurseries and garden centres and surrounding natural environments linked to major river systems |
List of species for which predictive models were developed, the bioclimatic predictors used to develop each model, and the percentage contribution of each predictor to the model.
| Species | Bioclimatic Predictors selected (% contribution to model) | Model Performance (AUC ± Standard Deviation) |
|---|---|---|
| Alternanthera philoxeroides | Mean diurnal range (10), Mean temperature of the warmest month (17), Precipitation seasonality (21), Precipitation of the warmest quarter (9), Precipitation of the coldest quarter (54) | High accuracy (0.929 ± 0.007) |
| Cenchrus echinatus | Mean temperature of the warmest quarter (25), Precipitation of seasonality (34), Precipitation of the wettest quarter (44), Precipitation of the driest quarter (7) | Moderate accuracy (0.812 ± 0.008) |
| Lithobates catesbeianus | Mean diurnal range (4), Temperature seasonality (44), Maximum temperature of the warmest month (21), Precipitation of the warmest quarter (3), Precipitation of the coldest quarter (38) | Moderate accuracy (0.791 ± 0.005) |
| Solenopsis invicta | Mean diurnal range (13), Maximum temperature of the warmest month (28), Precipitation of the wettest month (20), Precipitation of the driest month (45), Precipitation seasonality (4) | High accuracy (0.961 ± 0.006) |
Discussion
While watch lists and prohibited lists are beneficial in highlighting species to monitor, the lists often consist of numerous species, across a variety of taxa (e.g. the NEMBA prohibited species list – 553 targeted species;
The Convention on Biological Diversity (CBD) Aichi Target 9 requires that pathways of introduction be identified and prioritised for management efforts (UNEP, 2011). In this study, we identified likely sites of first naturalisation as priorities for incursion response efforts. We identified three important potential introduction points: the Durban Harbour; pet and aquarium stores; and nursery and garden centres. Each of the species used in this study were linked to one of these potential introduction points. The potential sites of first naturalisation identified in this study were all found to be in close proximity to the Durban harbour and the major river systems in the city, indicating that these sites are important for monitoring efforts.
Identifying the pathways facilitating the introduction of alien species is important for preventing alien species introductions. However, not all pathways of introduction are operational in all cities. By identifying the pathways which facilitate alien species introductions, priorities can be assigned to species with the potential of being introduced to the particular region of interest. In this study we were able to eliminate the species Vulpes vulpes (red fox) because the pathways facilitating its introduction (hunting in the wild and fur farms) are not operational in eThekwini. By contrast, the pathways which facilitate the introduction of C. echinatus are unknown. Therefore, determining if, how, and where the species is likely to be introduced to the city should be a key area for future applied research.
The Durban Harbour was identified as an important potential introduction point as well as a site to monitor for the introduction of A. philoxeroides and S. invicta. The pathways facilitating the introduction of these species are linked to the harbour. Alternanthera philoxeroides is primarily introduced through ship ballast and as a stowaway on ship cargo (
The river systems adjacent to potential point of introduction in the municipality were also identified as important sites to monitor. Alternanthera philoxeroides (
Invasions are, of course, often unpredictable and context dependent. Therefore the prioritisation here should only be one small part of an overall biosecurity strategy (
Even though this study focuses on eThekwini, the procedures used here represent a practical method to assign priorities for preventing the introduction of alien species. The methodology used in this study has merit for assigning priorities to a variety of taxa, such as this study (invertebrates, plants and vertebrates), or single taxa studies. Online databases such as CABI ISC, GBIF, GISD and GRIIS make alien species information required for utilising this methodology readily accessible. The accessibility of information and adaptability of the methodology used in this study makes the protocol feasible. However, there are many ways in which the protocol can be improved. For example, occurrence data sourced from online databases are often plagued with inconsistencies (e.g. validity of location points and taxonomy). The use of expert opinion in determining the validity of these data is a potentially beneficial improvement to this prioritisation tool. The procedures used in this study can further be improved quantitatively through additional analyses which will assess how pathways of introduction contribute to invasiveness (e.g. frequency analysis tests) of the target species as well as the contribution of potential introduction points to invasiveness (e.g. landscape level analysis) of target species. The advantage of the technique presented here is that it focuses on likely known threats and ensures that appropriate measures are put in place to deal with them.
Conclusion
Prioritisation is a fundamental component of effective strategic prevention strategies targeting the introduction of alien species to new regions (
Acknowledgements
This research was funded by the South African National Department of Environmental Affairs through its funding of the South African National Biodiversity Institute, Biological Invasions Directorate.
References
- Akmentins MS, Cardozo DE (2010) American bullfrog Lithobates catesbeianus (Shaw, 1802) invasion in Argentina. Biological Invasions 12: 735–737. https://doi.org/10.1007/s10530-009-9515-3
- Allen CR, Epperson DM, Garmestani AS (2004) Red imported fire ant impact on wildlife: A decade of research. The American Midland Naturalist 152: 88–103. https://doi.org/10.1674/0003 0031(2004)152[0088:RIFAIO]2.0.CO;2
- Ascunce MS, Yang CC, Oakey J, Calcaterra L, Wu WJ, Shih CJ, Goudet J, Ross KG, Shoemaker D (2011) Global invasion history of the fire ant Solenopsis invicta. Science 331: 1066–1068. https://doi.org/10.1126/science.1198734
- Austin MP (2002) Spatial prediction of species distribution: an interface between ecology theory and statistical modelling. Ecological Modelling 157: 101–118. https://doi.org/10.1016/S0304-3800(02)00205-3
- Barbet‐Massin M, Jiguet F, Albert CH, Thuiller W (2012) Selecting pseudo-absences for species distribution models: how, where and how many? Methods in Ecology and Evolution 3: 327–338. https://doi.org/10.1111/j.2041-210X.2011.00172.x
- Bassett IE, Beggs JR, Paynter Q (2010) Decomposition dynamics of invasive alligator weed compared with native sedges in a Northland lake. New Zealand Journal of Ecology 34: 324–331. https://newzealandecology.org/nzje/2940
- Branquart E (2007) Guidelines for environmental impact assessment and list classification of non-native organisms in Belgium. Version 2.4. Harmonia. Belgium Forum on Invasive Species. https://ias/biodiversity.be/ias/documents/ISEIA_protocol.pdf
- Brown J (2014) SDMtoolbox: a python-based GIS toolkit for landscape genetic, biogeographic and species distribution model analyses. Methods in Ecology and Evolution 5: 694–700. https://doi.org/10.1111/2041-210X.12200
- Burgin S, Norris A (2008) Alligator weed (Alternanthera philoxeroides) in New South Wales, Australia: A status report. Weed Biology and Management 8: 284–290. https://doi.org/10.1111/j.1445-6664.2008.00306.x
- Burgin S, Norris A, Karlson D (2010) Alternanthera philoxeroides in New South Wales, Australia: Are we closer to control of alligator weed? Weed Technology 24: 121–126. https://doi.org/10.1614/WT-08-059.1
- CABI (2018) Invasive Species Compendium. CAB International. http://www.cabi.org/isc [16 Feb 2018]
- Cadotte MW, Yasui SL, Livingstone S, MacIvor JS (2017) Are urban systems beneficial, detrimental, or indifferent for biological invasions? Biological Invasions 19: 3489–3470. https://doi.org/10.1007/s10530-017-1586-y
- Carpio AJ, Barasona JA, Guerrero‐Casado J, Oteros J, Tortosa FS, Acevedo P (2016) An assessment of conflict areas between alien and native species richness of terrestrial vertebrates on a macro-ecological scale in a Mediterranean hotspot. Animal Conservation 20: 433–443. https://doi.org/10.1111/acv.12330
- Chen Y, Zhou Y, Yin TF, Liu CX, Luo FL (2013) The invasive wetland plant Alternanthera philoxeroides shows a higher tolerance to waterlogging than its native congener Alternanthera sessilis. PLoS ONE 8: e81456. https://doi.org/10.1371/journal.pone.0081456
- Clements D, Dugdale TM, Hunt TD (2011) Growth of alligator weed (Alternanthera philoxeroides) over 5 years in South-east Australia. Aquatic Invasions 6: 77–88. https://doi.org/10.3391/ai.2011.6.1.09
- da Silva ET, Filho OPR (2009) Predation on juveniles of the invasive American bullfrog Lithobates catesbeianus (Anura: Ranidae) by native frog and snake species in South-eastern Brazil. Herpetology Notes 2: 215–218.
- da Silva ET, Dos Reis EP, Feio RN, Ribeiro Filho OP (2009) Diet of the invasive frog Lithobates catesbeianus (Shaw, 1802) (AnuraL Ranidae) in Viçosa, Minas Gerais State, Brazil. South American Journal of Herpetology 4: 286–294. https://doi.org/10.2994/057.004.0312
- DEA (2016) National environmental management: biodiversity act, 2004 (Act No. 10 of 2004) Alien and invasive species lists. Government Gazette 40166(864): 31–104.
- Dickie IA, Bennett BM, Burrows LE, Nunez MA, Peltzer DA, Porté A (2014) Conflicting values: Ecosystems services and invasive tree management. Biological Invasions 16: 705–719. https://doi.org/10.1007/s10530-013-0609-6
- Early R, Bradley BA, Dukes JS, Lawler JJ, Olden JD, Blumenthal DM, Gonzalez P, Grosholz ED, Ibañez I, Miller LP, Sorte CJ (2016) Global threats from invasive alien species in the twenty-first century and national response capacities. Nature Communications 7: 1–9. https://doi.org/10.1038/ncomms12485
- Elith J, Phillips SJ, Hastie T, Dudík M, Chee YE, Yates CJ (2011) A statistical explanation of MaxEnt for ecologists. Diversity and Distribution 17: 43–57. https://doi.org/10.1111/j.1472-4642.2010.00725.x
- Eskew EA, Worth SJ, Foley JE, Todd BD (2015) American bullfrogs (Lithobates catesbeianus) resists infection by multiple isolates of Batrachochytrium dendrobatidis including one implicated in wild mass mortality. Ecohealth 12: 513–518. https://doi.org/10.1007/s10393-015-1035-2
- ESRI (2015) ArcGIS ArcMap v10.3.1.4959. Environmental Systems Research Institute. Redlands, California, USA
- Essl F, Nehring S, Klingenstein F, Milasowszky N, Nowack C, Rabitsch W (2011) Review of risk assessment systems of IAS in Europe and introducing the German-Austrian Black List Information System (GABLIS). Journal of Nature Conservation 19: 339–350. https://doi.org/10.1016/j.jnc.2011.08.005
- Faulkner KT, Robertson MP, Rouget M, Wilson JR (2014) A simple, rapid methodology for developing invasive species watchlists. Biological Conservation 179: 25–32. https://doi.org/10.1016/j.biocon.2014.08.014
- Faulkner KT, Robertson MP, Rouget M, Wilson JR (2016a) Border control for stowaway alien species should be prioritised on variations in establishment debt. Journal of Environmental Management 108: 301–309. https://doi.org/10.1016/j.jenvman.2016.05.023
- Ficetola GF, Thuiller W, Miaud C (2007) Prediction and validation of the potential global distribution of a problematic alien invasive species – the American bullfrog. Diversity and Distribution 13: 467–485. https://doi.org/10.1111/j.1472-4642.2007.00377.x
- Gaertner M, Larson BM, Irlich UM, Holmes PM, Stafford L, van Wilgen BW, Richardson DM (2017) Managing invasive species in cities: A framework from Cape Town, South Africa. Landscape and Urban Planning 151: 1–9. https://doi.org/10.1016/j.landurbplan.2016.03.010
- Gaertner M, Wilson JR, Cadotte MW, MacIvor JS, Zenni RD, Richardson DM (2017) Non-native species in urban environments: patterns, processes, impacts and challenges. Biological Invasions 19: 3461–3470. https://doi.org/10.1007/s10530-017-1598-7
- GBIF (2017a) GBIF Occurrence download Alternanthera philoxeroides. https://doi.org/10.15468/dl.fay5de [Accessed 21 November 2017]
- GBIF (2017b) GBIF Occurrence download Cenchrus echinatus. https://doi.org/10.15468/dl.g5wqi0 [Accessed 21 November 2017]
- GBIF (2017c) GBIF Occurrence download Lithobates catesbeianus. https://doi.org/10.15468/dl.0xqd2c [Accessed 21 November 2017]
- GBIF (2017d) GBIF Occurrence download Solenopsis invicta. https://doi.org/10.15468/dl.i6nsbc [Accessed 21 November 2017]
- Genovesi P, Shine C (2004) European Strategy on Invasive Alien Species. Council of Europe (Nature and environment), No. 137, Council of Europe Publishing, Strasbourgh, 66 pp.
- GISD (2018) Global Invasive Species Database. Invasive Species Specialist Group (ISSG) of the IUCN Species Survival Commission. http://www.issg.org/database [16 Feb 2018]
- GRIIS (2017) Global Register of Introduced and Invasive Species. https://www.griis.org/ [05 May 2017]
- Hansen MJ, Clevenger AP (2005) The influence of disturbance and habitat on the presence of non-native plant species along transport corridors. Biological Conservation 125: 249–259. https://doi.org/10.1016/j.biocon.2005.03.024
- Harrower CA, Scalera R, Pagad S, Schonrogge K, Roy HE (2017) Guidance for interpretation of CBD categories on introduction pathways. Technical note prepared by IUCN for the European Commission.
- Hester SM, Cacho OJ (2017) The contribution of passive surveillance to invasive species management. Biological Invasions 19: 737–748. https://doi.org/10.1007/s10530-016-1362-4
- Hill MP, Clusella-Trullas S, Terblanche JS, Richardson DM (2016) Drivers, impacts, mechanisms and adaptations in insect invasions. Biological Invasions 18: 883–891. https://doi.org/10.1007/s10530-016-1088-3
- Hijmans RJ, Cameron SE, Parra JL, Jones PG, Jarvis A (2005) Very high resolution interpolated climate surfaces for global land areas. International Journal of Climatology 25: 1965–1978. https://doi.org/10.1002/joc.1276
- Holway DA, Lach L, Suarez AV, Tsutsui ND, Case TJ (2002) The causes and consequences of ant invasions. Annual Review of Ecology and Systematics 33: 181–233. https://doi.org/10.1146/annurev.ecolsys.33.010802.150444
- Hulme PE, Bacher S, Kenis M, Klotz S, Kühn I, Minchin D, Nentwig W, Olenin S, Panov V, Pergl J, Pyšek P (2008) Grasping at the routes of biological invasions: a framework for integrating pathways into policy. Journal of Applied Ecology 45: 403–414. https://doi.org/10.1111/j.1365-2664.2007.01442.x
- Hulme PE (2009) Trade, transport and trouble: managing invasive pathways in an era of globalization. Journal of Applied Ecology 46: 10–18. https://doi.org/10.1111/j.1365-2664.2008.01600.x
- Irlich UM, Potgieter L, Stafford L, Gaertner M (2017) Recommendations for municipalities to become compliant with National legislation on biological invasions. Bothalia: African Biodiversity and Conservation a2156. https://doi.org/10.4102/abc.v47i2.2156
- Julien MH, Skarratt B, Maywald GF (1995) Potential geographic distribution of Alligator weed and its biological control by Agasicles hygrophila. Journal of Aquatic Plant Management 33: 55–60.
- Kenis M, Auger-Rozenberg MA, Roques A, Timms L, Péré C, Cock MJ, Settele J, Augustin S, Lopez-Vaamonde C (2009) Ecological effects of invasive alien insects. Biological Invasions 11: 21–45. https://doi.org/10.1007/s10530-008-9318-y
- Kottek M, Grieser J, Beck C, Rudolf B, Rubel F (2006) World Map of Köppen-Geiger climate classification updated. Meteorologische Zeitschrift 15: 259–263. https://doi.org/10.1127/0941-2948/2006/0130
- Kowarik I (2011) Novel urban ecosystems, biodiversity, and conservation. Evironmental Pollution 159: 1974–1983. https://doi.org/10.1016/j.envpol.2011.02.022
- Kuhman TR, Pearson SM, Turner MG (2010) Effects of land-use history and the contemporary landscape on non-native plant invasions at local and regional scales in the forest dominated southern Appalachains. Landscape Ecology 25: 1433–1445. https://doi.org/10.1007/s10980-010-9500-3
- Lövei GL (1997) Biodiversity: global change through invasion. Nature 388: 627–628. https://doi.org/10.1038/41665
- Lowe S, Browne M, Boudjelas S, De Poorter M (2000) 100 of the World’s Worst Invasive Alien Species: A selection from the Global Invasive Species Database. The Invasive Species Specialist Group, New Zealand. https://portals.iucn.org/library/sites/library/files/documents/2000-126.pdf
- McGeoch MA, Genovesi P, Bellingham PJ, Costello MJ, McGrannachan C, Sheppard A (2016) Prioritizing species, pathways, and sites to achieve conservation targets for biological invasions. Biological Invasions 18: 299–314. https://doi.org/10.1007/s10530-015-1013-1
- McGlynn TP (1999) The worldwide transfer of ants: geographical distribution and ecological invasions. Journal of Biogeography 26: 535–548. https://doi.org/10.1046/j.1365-2699.1999.00310.x
- Measey J, Davies SJ, Vimercati G, Rebelo A, Schmidt W, Turner A (2017) Invasive amphibians in southern Africa: A review of invasion pathways. Bothalia 47: 1–12. https://doi.org/10.4102/abc.v47i2.2117
- Morrison LW, Porter SD, Daniels E, Korzukhin MD (2004) Potential global range expansion of the invasive fire ant (Solenopsis invicta). Biological Invasions 6: 183–191. https://doi.org/10.1023/B:BINV.0000022135.96042.90
- Nehring S, Klingenstein F (2008) Aquatic alien species in Germany – listing system and options for action. Neobiota 7: 19–33
- NEMBA (2014) National Environmental Management: Biodiversity Act 2004 (Act No. 10 of 2004) Alien and Invasive Species Lists 2014. Government Gazette No. 37886. Pretoria, South Africa.
- Padayachee AL, Irlich UM, Faulkner KT, Gaertner M, Procheş Ş, Wilson JR, Rouget M (2017) How do invasive species travel to and through the urban environment? Biological Invasions 19: 3557–3570. https://doi.org/10.1007/s10530-017-1596-9
- Parrott D, Roy S, Baker R, Cannon R, Eyre D, Hill M, Wagner M, Preston C, Roy H, Beckmann B, Copp GH (2009) Horizon scanning for new invasive non-native animal species in England. Natural England Commissioned Report NECR009, Sheffield.
- Pearson RG, Raxworthy CJ, Nakamura M, Townsend Peterson A (2007) Predicting species distributions from small numbers of occurrence records: a test case using cryptic geckos in Madagascar. Journal of Biogeography 34: 102–117. https://doi.org/10.1111/j.1365-2699.2006.01594.x
- Pergl J, Pysek P, Bacher S, Essl F, Genovesi P, Harrower CA, Hulme PE (2017) . Troubling travellers: are economically harmful alien species associated with particular introduction pathways? Neobiota 32: 1–20. https://doi.org/10.3897/neobiota.32.10199
- Peterson AT, Soberón J, Pearson RG, Anderson RP, Martínez-Meyer E, Nakamura M, Araújo MB (2011) Ecological niches and geographic distributions. Princeton University Press, Princeton, New Jersey. https://doi.org/10.23943/princeton/9780691136868.001.0001
- Phillips SJ, Anderson RP, Schapire RE (2006) Maximum entropy modelling of species geographic distribution. Ecological Modelling 190: 231–259. https://doi.org/10.1016/j.ecolmodel.2005.03.026
- Phillips SJ, Dudík M (2008) Modeling of species distributions with Maxent: new extensions and comprehensive evaluation. Ecography 31: 161–175. https://doi.org/10.1111/j.0906-7590.2008.5203.x
- Pimentel D, McNair S, Janecka J, Wightman J, Simmonds C, O’connell C, Wong E, Russel L, Zern J, Aquino T, Tsomondo T (2001) Economic and environmental threats of alien plants, animal and microbe invasions. Agricultural Ecosystems and Environments 84: 1–20. https://doi.org/10.1016/S0167-8809(00)00178-X
- Potgieter LJ, Gaertner M, Kueffer C, Larson BM, Livingstone SW, O’Farrell PJ, Richardson DM (2017) Alien plants as mediators of ecosystems and disservices in urban systems: a global review. Biological Invasions 19: 3571–3558. https://doi.org/10.1007/s10530-017-1589-8
- Pyšek P, Jarošík V, Hulme PE, Kühn I, Wild J, Arianoutsou M, Bacher S, Chiron F, Didžiulis V, Essl F, Genovesi P (2010) Disentangling the role of environmental and human pressures on biological invasions across Europe. Proceedings of the National Academy of Science 107: 12157–12162. https://doi.org/10.1073/pnas.1002314107
- Reaser JK, Meyerson LA, Von Holle B (2008) Saving camels from straws: how propagule pressure-based prevention policies can reduce the risk of biological invasions. Biological Invasions 10: 1085–1098. https://doi.org/10.1007/s10530-007-9186-x
- Rejmánek M, Richardson DM (1996) What attributes make some plant species more invasive? Ecology 77: 1655–1661. https://doi.org/10.2307/2265768
- Richardson DM, Pyšek P, Rejmánek M, Barbour MG, Panetta FD, West CJ (2000) Naturalization and invasion of alien plants: concepts and definitions. Diversity and Distributions 6: 93–107. https://doi.org/10.1046/j.1472-4642.2000.00083.x
- Roberts D (2008) Thinking globally, acting locally – institutionalizing climate change at the local government level in Durban, South Africa. Environment and Urbanization 20: 521–537. https://doi.org/10.1177/0956247808096126
- Robertson MP, Villet MH, Palmer AR (2004) A fuzzy classification technique for predicting species distributions: applications using invasive alien plants and indigenous insects. Diversity and Distribution 10: 461–474. https://doi.org/10.1111/j.1366-9516.2004.00108.x
- Scalera R, Genovesi P, Booy O, Essl F, Jeschke J, Hulme P, Wilson J (2016) Technical Report: Progress towards pathways prioritization in compliance to Aichi Target 9, Information documented presented at SBSTTA 20 UNEP/CBC/SBSTTA/20/INF/S, the twentieth meeting of the CBD’s Subsidiary Body on Scientific, Technical and Technological Advice, Montreal, Canada, 25–30 April 2016.
- Snedecor GW, Cochran WG (1968) Statistical Methods (6th edn). Ames Iowa, The Iowa State University Press.
- Solley GO, Vanderwoude C, Knight GK (2002) Anaphylaxis due to red imported fire ant sting. Medical Journal of Australia 176: 521–523.
- STATSSA (2017) Statistics South Africa. http://www.statssa.gov.za/?page_id=1021&id=ethekwini-municipality [15 Oct 2018]
- Tang L, Sun YY, Zhang QP, Zhou Y, Zhang N, Zhang ZX (2013) Fumigant activity of eight plant essential oils against workers of red imported fire ant, Solenopsis invicta. Sociobiology 60: 35–40. https://doi.org/10.13102/sociobiology.v60i1.35-40
- Tatem AJ (2009) The worldwide airline network and the dispersal of exotic species: 2007–2010. Ecography 32: 94–102. https://doi.org/10.1111/j.1600-0587.2008.05588.x
- Thuiller W, Richardson DM, Rouget M, Procheş Ş, Wilson JR (2006) Interactions between environment, species traits and human uses describe patterns of plant invasions. Ecology 87: 1755–1769. https://doi.org/10.1890/0012-9658(2006)87[1755:IBESTA]2.0.CO;2
- UNEP (2011) The strategic plan for biodiversity 2011–2020 and the Aichi biodiversity targets. COP CBD tenth meeting, Nagoya (Japan), October 2010. http://www.cbd.int/decisions/cop/?m=cop-10
- Verbrugge LNH, Lueven RSEW, van der Velde G (2010) Evaluation of international risk assessment protocols for exotic species. Final Report. University Nijmegen, Netherlands.
- Vilà M, Basnou C, Pyšek P, Josefsson M, Genovesi P, Gollasch S, Nentwig W, Olenin S, Roques A, Roy D, Hulme PE (2010) How well do we understand the impacts of alien species on ecosystem services? A pan-European, cross-taxa assessment. Frontiers in Ecology and Environment 8: 135–144. https://doi.org/10.1890/080083
- Vilà M, Espinar JL, Hejda M, Hulme PE, Jarošík V, Maron JL, Pergl J, Schaffner U, Sun Y, Pyšek P (2011) Ecological impacts of invasive alien plants: a meta-analysis of their effects on species, communities and ecosystems. Ecology Letters 14: 702–708. https://doi.org/10.1111/j.1461-0248.2011.01628.x
- Welk E, Schubert K, Hoffmann MH (2002) Present and potential distribution of invasive garlic mustard (Alliaria petiolata) in North America. Diversity and Distribution 8: 219–233. https://doi.org/10.1046/j.1472-4642.2002.00144.x
- Wilson JR, Panetta FD, Lindgren C (2017) Detecting and responding to alien plant incursions. Cambridge University Press – Ecology, Biodiversity and Conservation Series, 286 pp. https://doi.org/10.1017/CBO9781316155318
- Zengeya T, Ivey P, Woodford DJ, Weyl O, Novoa A, Shackleton R, Richardson D, Van Wilgen B (2017) Managing conflict-generating invasive species in South Africa: Challenges and trade-offs. Bothalia 47:a2160. https://doi.org/10.4102/abc.v47i2.2160