Data Paper |
|
Corresponding author: Jacob E. Lucero ( lucero.jacob.e@gmail.com ) Academic editor: Marcel Rejmanek
© 2019 Jacob E. Lucero, Taylor Noble, Stephanie Haas, Michael Westphal, H. Scott Butterfield, Christopher J. Lortie.
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Lucero JE, Noble T, Haas S, Westphal M, Butterfield HS, Lortie CJ (2019) The dark side of facilitation: native shrubs facilitate exotic annuals more strongly than native annuals. NeoBiota 44: 75-93. https://doi.org/10.3897/neobiota.44.33771
|
Abstract
Positive interactions enhance biodiversity and ecosystem function, but can also exacerbate biological invasions. Facilitation of exotic invaders by exotic foundation species (invasional meltdown) has been studied extensively, but facilitation of exotic invaders by native foundation species has attracted less attention. Specifically, very few studies have examined the extent that native foundation species facilitate native and exotic competitors. Understanding the processes that mediate interactions between native and exotic species can help explain, predict, and improve management of biological invasions. Here, we examined the effects of native foundation shrubs on the relative abundance of the annual plant community – including native and exotic taxa – from 2015–2018 in a desert ecosystem at Carrizo Plain National Monument, California, USA (elevation: 723 m). Shrub effects varied by year and by the identity of annual species, but shrubs consistently enhanced the abundance of the annual plant community and facilitated both native (n=17 species) and exotic (n=4 species) taxa. However, at the provenance level, exotic annuals were facilitated 2.75 times stronger in abundance than native annuals, and exotic annuals were always more abundant than natives both near and away from shrubs. Our study reaffirms facilitation as an important process in the organisation of plant communities and confirms that both native and exotic species can form positive associations with native foundation species. However, facilitation by native foundation species can exacerbate biological invasions by increasing the local abundance of exotic invaders. Thus, the force of facilitation can have a dark side relevant to ecosystem function and management.
Keywords
Bromus rubens, deserts, exotic species, facilitation, invasional meltdown, native species, plant invasions, shrubs
Introduction
Positive interactions among species, or facilitation, can strongly influence the organisation of plant communities (
However, facilitation can have a dark side when beneficiary species are exotic invaders. Invasive plant species pose a pervasive threat to ecosystem function worldwide (
The objective of this study was to investigate the extent that native and exotic species of annual plants associate with (i.e., are facilitated by) native foundation shrubs in an arid ecosystem. This issue is timely because drylands worldwide are increasingly comprised of exotic species (
Methods
Study site and species
We surveyed annual plant communities at Carrizo Plain National Monument in the San Joaquin Desert (
Sampling
We sampled the abundance of the annual plant community using a paired shrub-open microsite contrast with 0.5 × 0.5 m quadrats (
Statistical analyses
Relative interaction indices (“RIIs” hereafter;
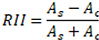
where As is the abundance (i.e., no. individual plants) of an annual species in a shrub microsite and Ac is the abundance of the same annual species in the paired open microsite. RII values range from -1 to +1. Negative RII values indicate negative (competitive) effects of shrubs on annual species, positive values indicate positive (facilitative) effects, and a value of 0 indicates a neutral effect. Annuals are only considered to be beneficiary species of foundation shrubs when RII is positive.
To evaluate whether annual species were generally more abundant near E. californica shrubs than away from shrubs, we performed independent one-sample t-tests with mean RII (pooled per species per year) as the response variable for each year of the study. We used an additional one-sample t-test to summarise the net strength of shrub associations across all study years. We evaluated the net strength of species-specific shrub associations using a linear mixed-effects model with RII (pooled per species per year) as the response variable, species identity and shrub volume as fixed factors, and study year as a random factor. Treating year as a random factor accounted for stochastic sources of inter-annual variation, such as climate (Suppl. material
To test whether native and exotic annual species associated differentially with E. californica at the provenance level, we employed independent generalised linear models for each year of the study with RII (pooled per species) as the response variable and species provenance as a fixed factor. We contrasted the net strength of native vs. exotic associations with shrubs across all study years using a linear mixed-effects model with RII (pooled per species per year) as the response variable, species provenance and shrub volume as fixed factors, and study year as a random factor.
We inferred the outcome of biotic interactions between native and exotic annuals at the provenance level using t-tests and linear models. For shrub and open microsites and for each year of the study, we contrasted the net abundance of native vs. exotic annuals using independent one-sample t-tests with abundance (i.e., not RIIs) as the response variable. We then regressed net native abundance against net exotic abundance using linear models. These regressions addressed the effects of exotic annual species on the abundance of native annual species. Negative line slopes suggested competitive effects; positive line slopes suggested facilitative effects (
All analyses were performed in R, version 3.5.1 (
Results
Annual plant species were generally more abundant near native foundation shrubs than away from shrubs. Across all species and years, annual plants were 1.35 (± 0.68 SE) times more abundant under shrubs than in the open (df = 2381, t-value = 12.97, P < 0.01). Accordingly, net RII summarised across all species and years was greater than zero (RII = 0.22 ± 0.05 SE, df = 30.00, t-value = 4.98, P << 0.01) (Table
Mean effects (RII ± SE) of shrubs on annual species at the community level. RII values are summarised across all annual species for each year of the study (2015–2018) and for all study years combined (Net). P ≤ 0.05 indicates that RII values differ significantly from zero, according to independent t-tests.
| Year | RII. | df | t-value | P-value |
|---|---|---|---|---|
| 2015 | 0.18 (0.07) | 11.00 | 2.86 | 0.01 |
| 2016 | 0.10 (0.05) | 8.00 | 2.21 | 0.05 |
| 2017 | 0.39 (0.31) | 3.00 | 2.06 | 0.13 |
| 2018 | 0.38 (0.14) | 5.00 | 3.62 | 0.01 |
| Net | 0.22 (0.05) | 30.0 | 4.98 | <<0.01 |
In each year of the study, native and exotic annual species positively associated with E. californica to become beneficiary species (Fig.

Year-by-year effects (RII ± 95% CI) of native foundation shrubs on the abundance of annual species. Net effects across all years are summarised in Suppl. material
Interestingly, native and exotic annuals associated differentially with native foundation shrubs at the provenance level. At the provenance level, net RII summarised across all years was 2.75 ± 0.14 SE times greater for exotic annuals than for native annuals (df = 21.01, Z-ratio = 3.05, P < 0.01), and this general trend (i.e., greater RII values for exotic annuals than native annuals at the provenance level) was apparent in each year of the study except 2017 (Table
Mean effects (RII ± SE) of shrubs on annual species at the provenance level. RII values are summarised across native (RIInative) and exotic (RIIexotic) annual species for each year of the study (2015–2018) and for all study years combined (Net). P ≤ 0.05 indicates that RII values differ at the provenance level, according to independent generalized linear models (2015–2018) and a linear mixed-effects model (Net).
| Year | RIInative | RIIexotic | df | Z-ratio | P-value |
|---|---|---|---|---|---|
| 2015 | 0.13 (0.06) | 0.41 (0.14) | 11.00 | 1.90 | 0.05 |
| 2016 | 0.05 (0.05) | 0.21 (0.07) | 8.00 | 1.91 | 0.05 |
| 2017 | 0.25 (0.18) | 0.82 (0.30) | 3.00 | 1.61 | 0.10 |
| 2018 | 0.24 (0.09) | 0.64 (0.12) | 5.00 | 2.66 | <0.01 |
| Net | 0.16 (0.13) | 0.44 (0.15) | 21.01 | *9.28 | <0.01 |
Regardless of year and microsite, exotic annual species were always more abundant than native annual species. Summarised across all years, the net abundance of exotic annuals was 4.97 ± 0.78 SE and 3.05 ± 0.78 SE times greater than the net abundance of native annuals in shrub and open microsites, respectively (df ≥ 1863.40, t-value ≥ |28.89|, P < 0.01) (Suppl. material
At the provenance level, the relationship between the abundance of native and exotic annuals varied considerably by year (Fig.

Discussion
Facilitation is an important process in the assembly of plant communities in drylands and other extreme environments globally (
Our study coincides with a broad literature suggesting that ecological processes can have markedly different effects on native and exotic taxa in the same communities (reviewed by
Our study underscores the potential for facilitation by native foundation species to exacerbate biological invasions. Native foundation species can increase the ecophysiological performance (
There was considerable inter-annual variation in the effects of E. californica on annual species. The strength of positive interactions is known to increase with environmental severity (
We observed no effects of shrub size on association patterns at either the species or provenance levels (Suppl. material
Our study hints that facilitation by E. californica shrubs can alter the outcome of interspecific interactions among native and exotic neighbours. In 2018, the abundance of native and exotic annuals was positively related in open microsites (where E. californica was absent) but unrelated in shrub microsites (where E. californica was present). These relationships suggest facilitation between native and exotic annuals in open but not shrub microsites. Thus, it is possible that E. californica attenuated positive interactions between native and exotic annuals in that year. Facilitation of native species by exotic neighbours – including invasive species – is not necessarily unusual in deserts. For instance, in the Great Basin Desert,
Our study highlights the potential for beneficiary species to experience facilitation from foundation species and interspecific competition from other beneficiary species simultaneously. In 2015, competition and facilitation appeared to operate in tandem to influence biodiversity patterns under shrubs (Fig.
We hypothesised that any competitive effects of exotic annuals on native annuals in 2015 may have been driven by B. rubens, as this exotic species was facilitated more strongly and consistently than any other (Fig.
Our data do not speak to the mechanisms by which facilitation occurred. Non-mutually exclusive mechanisms of facilitation include seed trapping, amelioration of abiotic stress, modification of soil biogeochemical processes, increasing pollinator visitation, and/or providing herbivore protection (reviewed by
Our findings have practical implications. Because E. californica canopies were hotspots for the abundance of native and exotic annual species, conservationists may consider targeting their efforts to control invasive species under shrub canopies. For example, herbicide applications to reduce the density of invasive species and subsequent reseeding efforts to increase the density of native species (
Furthermore, our study suggests caution in using facilitation by native shrubs as a tool for restoring native biodiversity to degraded environments. Drylands in California and globally are being retired from intensive agricultural use due to drought, poor soils, and changing climate (
Conclusions
Our study reaffirms facilitation as an important force in the organisation of plant communities and confirms that both native and exotic beneficiary species can positively associate with native foundation shrubs. However, we found that the magnitude of facilitation depended upon the biogeographic origins of beneficiary species – at the provenance level, exotic species were facilitated in abundance much stronger than native species. Importantly and regardless of inter-annual variation in climate, the net outcome of biotic interactions that included facilitation was an annual plant community dominated (in terms of relative abundance) by exotic species. Our study stresses that the effects of ecological processes like facilitation must not be decoupled from net outcomes relevant to conservation and restoration. In systems like ours where facilitation increases the abundance of invasive species, managing positive interactions may be a useful conservation strategy.
Acknowledgements
This research was funded by a TNC grant, BLM funds, and an NSERC DG to CJL. JEL was supported by a York Science Fellowship. Ray Callaway provided excellent editorial input.
References
- Allington GRH, Koons DN, Morgan Ernest SK, Schutzenhofer MR, Valone TJ (2013) Niche opportunities and invasion dynamics in a desert annual community. Ecology Letters 16: 158–166. https://doi.org/10.1111/ele.12023
- Altieri AH, van Wesenbeeck BK, Bertness MD, Silliman BR (2010) Facilitation cascade drives positive relationship between native biodiversity and invasion success. Ecology 91: 1269–1275. https://doi.org/10.1890/09-1301.1
- Armas C, Ordinales R, Pugnaire FI (2004) Measuring plant interactions: a new comparative index. Ecology 85: 2682–2686. https://doi.org/10.1890/03-0650
- Badano EI, Bustamante RO, Villarroel E, Marquet PA, Cavieres LA (2015) Facilitation by nurse plants regulates community invasibility in harsh environments. Journal of Vegetation Science 26: 756–767. https://doi.org/10.1111/jvs.12274
- Balch JK, Bradley BA, D’Antonio CM, Gomez-Dans J (2013) Introduced annual grass increases regional fire activity across the arid western USA (1980–2009). Global Change Biology 19: 173–183. https://doi.org/10.1111/gcb.12046
- Baxter CV, Fausch KD, Murakami M, Chapman PL (2004) Fish invasion restructures stream and forest food webs by interrupting reciprocal prey subsidies. Ecology 85: 2656–2663. https://doi.org/10.1890/04-138
- Bellard C, Cassey P, Blackburn TM (2016) Alien species as a driver of recent extinctions. Biology Letters 12. https://doi.org/10.1098/rsbl.2015.0623
- Bertness MD, Callaway RM (1994) Positive interactions in communities. Trends in Ecology and Evolution 9: 191–193. https://doi.org/10.1016/0169-5347(94)90088-4
- Blackburn TM, Pysek P, Bacher S, Carlton JT, Duncan RP, Jarosik V, Wilson JRU, Richardson DM (2011) A proposed unified framework for biological invasions. Trends in Ecology and Evolution 26: 333–339. https://doi.org/10.1016/j.tree.2011.03.023
- Brathen KA, Lortie CJ (2015) A portfolio effect of shrub canopy height on species richness in both stressful and competitive environments. Functional Ecology 30: 60–69. https://doi.org/10.1111/1365-2435.12458
- Brooker RW, Maestre FT, Callaway RM, Lortie CL, Cavieres LA, Kunstler G, Linacourt P, Tielborger K, Travis JM, Anthelme F, Armas C, Coll L, Corcket E, Delzon S, Forey E, Kikvidze Z, Olofsson J, Pugnaire F, Quiroz C, Saccone P, Schiffers K, Seifan M, Touzard B, Micharet R (2008) Facilitation in plant communities: the past, the present, and the future. Journal of Ecology 96: 18–34.
- Brooker R, Kikvidze Z, Pugnaire FI, Callaway RM, Choler P, Lortie CJ, Michalet R (2005) The importance of importance. Oikos 109: 63–70. https://doi.org/10.1111/j.0030-1299.2005.13557.x
- Brooks ML (1999) Alien annual grasses and fire in the Mojave Desert. Madroño 46: 13–19.
- Brooks ML (2000) Competition between alien annual grasses and native annual plants in the Mojave Desert. The American Midland Naturalist 144: 92–108. https://doi.org/10.1674/0003-0031(2000)144[0092:CBAAGA]2.0.CO;2
- Brooks ML, D’Antonio CM, Richardson DM, Grace JB, Keeley JE, DiTomaso JM, Hobbs RJ, Pellant M, Pyke D (2004) Effects of invasive alien plants on fire regimes. BioScience 54: 677–688. https://doi.org/10.1641/0006-3568(2004)054[0677:EOIAPO]2.0.CO;2
- Buck-Diaz J, Evens J (2011) Carrizo Plain National Monument vegetation classification and mapping project.
- Butterfield BJ, Cavieres LA, Callaway RM, Cook J, Kikvidze Z, Lortie CJ, Michalet R, Pugnaire FI, Schob C, Xiao S, Zaitchek B, Anthelme F, Bjork RG, Dickinson K, Gavilan R, Kanka R, Maalouf JP, Noroozi J, Parajuli R, Phoenix GK, Reid A, Ridenour W, Rixen C, Wipf S, Zhao L, Brooker RW (2013) Alpine cushion plants inhibit the loss of phylogenetic diversity in severe environments. Ecology Letters 16: 478–486. https://doi.org/10.1111/ele.12070
- Calflora (2018) Information on California plants for education, research and conservation, with data contributed by public and private institutions and individuals, including the Consortium of California Herbaria. Berkeley, California. https://www.calflora.org/ [Accessed: September 20, 2018]
- Callaway RM (1995) Positive interactions among plants. The Botanical Review 61: 306–349. https://doi.org/10.1007/BF02912621
- Callaway RM, DeLucia EH, Moore D, Nowak R, Schlesinger WH (1996) Competition and facilitation: contrasting effects of Artemisia tridentata on desert vs. montane pines. Ecology 77: 2130–2141. https://doi.org/10.2307/2265707
- Callaway RM (2007) Positive interactions and interdependence in plant communities. Springer, Dordrecht, The Netherlands.
- Callaway RM, Waller LP, Diaconu A, Pal R, Collins AR, Mueller-Schaerer H, Maron JL (2011) Escape from competition: neighbors reduce Centaurea stoebe performance at home but not away. Ecology 92: 2208–2213. https://doi.org/10.1890/11-0518.1
- Catford JA, Jansson R, Nilsson C (2009) Reducing redundancy in invasion ecology by integrating hypotheses into a single theoretical framework. Diversity and Distributions 15: 22–40. https://doi.org/10.1111/j.1472-4642.2008.00521.x
- Cavieres LA, Quiroz CL, Molina-Montenegro MA (2008) Facilitation of the non-native Taraxacum officinale by native nurse cushion species in the high Andes of central Chile: are there differences between nurses? Functional Ecology 22: 148–156.
- Cook BI, Ault TR, Smerdon JE (2015) Unprecedented 21st century drought risk in the American Southwest and Central Plains. Science Advances 1: e1400082. https://doi.org/10.1126/sciadv.1400082
- Clements CD, Harmon DN, Blank RR, Weltz M (2017) Improving seeding success on cheatgrass-infested rangelands in northern Nevada. Rangelands 39: 174–181. https://doi.org/10.1016/j.rala.2017.10.003
- Cutlar HC (1939) Monograph of the North American species of the genus Ephedra. Annals of the Missouri Botanical Garden 26: 373–428. https://doi.org/10.2307/2394299
- D’Antonio CM, Vitousek PM (1992) Biological invasions by exotic grasses, the grass/fire cycle, and global change. Annual Review of Ecology and Systematics 23: 63–87. https://doi.org/10.1146/annurev.es.23.110192.000431
- Davis MH et al. (2011) Don’t judge species on their origins. Nature 474: 153–154. https://doi.org/10.1038/474153a
- Fagundes M, Weisser W, Ganade G (2018) The role of nurse successional stages on species- specific facilitation in drylands: nurse traits and facilitation skills. Ecology and Evolution 8: 5173–5184. https://doi.org/10.1002/ece3.3962
- Filazzola A, Lortie CJ (2014) A systematic review and conceptual framework for the mechanistic pathways of nurse plants. Global Ecology and Biogeography 23: 1335–1345. https://doi.org/10.1111/geb.12202
- Filazzola A, Westphal M, Powers M, Liczner AR, (Smith) Woollett DA, Johnson B, Lortie CJ (2017) Non-trophic interactions in deserts: facilitation, interference, and an endangered lizard species. Basic and Applied Ecology 20: 51–61. https://doi.org/10.1016/j.baae.2017.01.002
- Filazzola A, Sotomayor DA, Lortie CJ (2018) Modelling the niche space of desert annuals to include positive interactions. Oikos 127: 264–273. https://doi.org/10.1111/oik.04688
- Foronda A, Pueyo Y, Arroyo AI, Saiz H, de la Luz Giner M, Alados CL (2019) The role of nurse shrubs on the spatial patterning of plant establishment in semi-arid gypsum plant communities. Journal of Arid Environments 160: 82–90. https://doi.org/10.1016/j.jaridenv.2018.09.003
- Freeman ED, Sharp TR, Larsen RT, Knight RN, Slater SJ, McMillan BR (2014) Negative effects of an exotic grass invasion on small-mammal communities. PLoS ONE 9: e108843. https://doi.org/10.1371/journal.pone.0108843
- Fridley JD, Stachowicz JJ, Naeem S, Sax DF, Seabloom EW, Smith MD, Stohlgren TJ, Tilman D, Von Holle B (2007) The invasion paradox: reconciling pattern and process in species invasions. Ecology 88: 3–7. https://doi.org/10.1890/0012-9658(2007)88[3:TIPRPA]2.0.CO;2
- Funk JL, Cleland EE, Suding KN, Zavaleta ES (2008) Restoration through reassembly: plant traits and invasion resistance. Trends in Ecology and Evolution 23: 695–703. https://doi.org/10.1016/j.tree.2008.07.013
- Gao X, Liu Z, Zhao X, Ling Q, Huo G, Wu P (2018) Extreme natural drought enhances interspecific facilitation in semiarid agroforestry systems. Agriculture, Ecosystems & Environment 265: 444–453. https://doi.org/10.1016/j.agee.2018.07.001
- Germano DJ, Rathburn GB, Saslaw LR, Cypher BL, Cypher EA, Vredenburgh LM (2011) The San Joaquin Desert of California: ecologically misunderstood and overlooked. Natural Areas Journal 31: 138–147. https://doi.org/10.3375/043.031.0206
- Gomez-Aparicio L (2009) The role of plant interactions in the restoration of degraded ecosystems: a meta-analysis across life-forms and ecosystems. Journal of Ecology 97: 1202–1214. https://doi.org/10.1111/j.1365-2745.2009.01573.x
- Griffith AB (2010) Positive effects of native shrubs on Bromus tectorum demography. Ecology 91: 141–154. https://doi.org/10.1890/08-1446.1
- He Q, Bertness MD, Altieri AH (2013) Global shifts towards positive species interactions with increasing environmental stress. Ecology Letters 16: 695–706. https://doi.org/10.1111/ele.12080
- Horn KJ, St. Clair SB (2017) Wildfire and exotic grass invasion alter plant productivity in response to climate variability in the Mojave Desert. Landscape Ecology 32: 635–646. https://doi.org/10.1007/s10980-016-0466-7
- Huddleston RT, Young TP (2005) Weed control and soil amendment effects on restoration plantings in an Oregon grassland. Western North American Naturalist 65: 507–515.
- Hupp N, Llambi LD, Ramírez L, Callaway RM (2017) Alpine cushion plants have species- specific effects on microhabitat and community structure in the tropical Andes. Journal of Vegetation Science 28: 928–938. https://doi.org/10.1111/jvs.12553
- Jeschke JM, Aparicio LG, Haider S, Heger T, Lortie CJ, Pysek P, Strayer D (2012) Support for major hypotheses in invasion biology is uneven and declining. NeoBiota 14: 1–20. https://doi.org/10.3897/neobiota.14.3435
- Kelsey R, Hart A, Butterfield HS, Vink D (2018) Groundwater sustainability in the San Joaquin Valley: multiple benefits if agricultural lands are retired and restored strategically. California Agriculture 72: 151–154. https://doi.org/10.3733/ca.2018a0029
- Kuznetsova A, Brockhoff PB, Christensen RHB (2017) lmerTest Package: Tests in Linear Mixed Effects Models. Journal of Statistical Software 82: 1–26. https://doi.org/10.18637/jss.v082.i13
- Lenth R, Singmann H, Love J, Buerkner P, Herve M (2018) Package ‘emmeans’. https://github.com/rvlenth/emmeans
- Lenz TI, Facelli JM (2003) Shade facilitates an invasive stem succulent in a chenopod shrubland in South Australia. Austral Ecology 28: 480–490. https://doi.org/10.1046/j.1442-9993.2003.01304.x
- Levine JM, Vila M, D’Antonio CM, Dukes JD, Grigulis K, Lavorel S (2003) Mechanisms underlying the impacts of exotic plant invasions. Proceedings of the Royal Society B 270: 775–781. https://doi.org/10.1098/rspb.2003.2327
- Liao C, Peng R, Lup Y, Zhou X, Wu X, Fang C, Chen J, Li B (2008) Altered ecosystem carbon and nitrogen cycles by plant invasion: a meta-analysis. New Phytologist 177: 706–714. https://doi.org/10.1111/j.1469-8137.2007.02290.x
- Llambi LD, Hupp N, Saez A, Callaway RM (2018) Reciprocal interactions between a facilitator, natives, and exotics in tropical alpine plant communities. Perspectives in Plant Ecology, Evolution, and Systematics. 30: 82–88. https://doi.org/10.1016/j.ppees.2017.05.002
- Lortie CJ (2018) Carrizo long-term ecological data analyses. Zenodo.
- Lortie CJ, Gruber E, Filazzola A, Noble T, Westphal M (2018a) The Groot effect: plant facilitation and desert shrub regrowth following extensive damage. Ecology and Evolution 8: 706–715. https://doi.org/10.1002/ece3.3671
- Lortie CJ, Noble T, Haas S, Butterfield S, Lucero J, Westphal M (2018b) Long-term ecological vegetation data for Carrizo National Monument, California. Knowledge Network for Biocomplexity.
- Lortie CJ, Filazzola A, Kelsey R, Hart AK, Butterfield S (2018c) Better late than never: a synthesis of strategic land retirement and restoration in California. Ecosphere 9: e02367. https://doi.org/10.1002/ecs2.2367
- Lucero JE, Allen PS, McMillan BR (2015) Increased primary production from an exotic invader does not subsidize native rodents. PLoS ONE 10(8): e0131564. https://doi.org/10.1371/journal.pone.0131564
- Lucero JE (2018) Do seeds from invasive bromes experience less granivory than seeds from native congeners in the Great Basin Desert? Plant Ecology 219: 1053–1061. https://doi.org/10.1007/s11258-018-0858-7
- Lucero JE, Callaway RM (2018) Native granivores reduce the establishment of native grasses but not invasive Bromus tectorum. Biological Invasions 20: 3491–3497. https://doi.org/10.1007/s10530-018-1789-x
- Maestre FT, Callaway RM, Valladares F, Lortie CJ (2009) Refining the stress-gradient hypothesis for competition and facilitation in plant communities. Journal of Ecology 2009: 199–205. https://doi.org/10.1111/j.1365-2745.2008.01476.x
- Maestre FT, Cortina J, Bautista S (2004) Mechanisms underlying the interaction between Pinus halipensis and the native late-successional shrub Pistacia lentiscus in a semi-arid plantation. Ecography 27: 776–786. https://doi.org/10.1111/j.0906-7590.2004.03990.x
- Maron JL, Vila M (2001) When do herbivores affect plant invasion? Evidence for the natural enemies and biotic resistance hypotheses. Oikos 95: 361–373. https://doi.org/10.1034/j.1600-0706.2001.950301.x
- Maron JL, Pearson DE, Potter T, Ortega YK (2012) Seed size and provenance mediate the joint effects of disturbance and seed predation on community assembly. Journal of Ecology 100: 1492–1500. https://doi.org/10.1111/j.1365-2745.2012.02027.x
- McIntire EJB, Fajardo A (2014) Facilitation as a ubiquitous driver of biodiversity. New Phytologist 201: 403–416. https://doi.org/10.1111/nph.12478
- Michalet R (2006) Is facilitation in arid environments the result of direct or complex interactions? New Phytologist 169: 3–6. https://doi.org/10.1111/j.1468-8137.2006.01617.x
- Michalet R, Pugnaire FI (2016) Facilitation in communities: underlying mechanisms, community and ecosystem implications. Functional Ecology 30: 3–9. https://doi.org/10.1111/1365-2435.12602
- Miriti LM (2006) Ontogenetic shift from facilitation to competition in a desert shrub. Journal of Ecology 94: 973–979. https://doi.org/10.1111/j.1365-2745.2006.01138.x
- Mitchell CE, Agrawal AA, Bever JD, Gilbert GS, Hufbauer RA, Klironomos JN, Maron JL, Morris WF, Parker IM, Power AG, Seabloom EW, Torchin ME, Vazquez DP (2006) Biotic interactions and plant invasions. Ecology letters 9: 726–740. https://doi.org/10.1111/j.1461-0248.2006.00908.x
- Noble TJ, Lortie CJ, Westphal M, Butterfield HS (2016) A picture is worth a thousand data points: an imagery dataset of paired shrub-open microsites within Carrizo Plain National Monument. GigaScience 5. https://doi.org/10.1186/s13742-016-0145-2
- Noumi Z, Chaieb M, Michalet R, Touzard B (2015) Limitations to the use of facilitation as a restoration tool in arid grazed savanna: a case study. Applied Vegetation Science 18: 391–401. https://doi.org/10.1111/avsc.12158
- Ostoja SM, Schupp EW (2009) Conversion of sagebrush shrublands to exotic annual grasslands negatively impacts small mammal communities. Diversity and Distributions 15: 863–870. https://doi.org/10.1111/j.1472-4642.2009.00593.x
- Padilla FM, Pugnaire FI (2006) The role of nurse plants in the restoration of degraded environments. Frontiers in Ecology and the Environment 4: 196–202. https://doi.org/10.1890/1540-9295(2006)004[0196:TRONPI]2.0.CO;2
- Pearson DE, Callaway RM (2008) Weed-biocontrol insects reduce native-plant recruitment through second-order apparent competition. Ecological Applications 18: 1489–1500. https://doi.org/10.1890/07-1789.1
- Pearson DE, Ortega YK, Ozkan E, Hierro JL (2016) Quantifying “apparent” impact and distinguishing impact from invasiveness in multispecies plant invasions. Ecological Applications 26: 162–173. https://doi.org/10.1890/14-2345
- Pearson DE, Ortega YK, Ozkan E, Hierro JL (2018) Community assembly theory as a framework for biological invasions. Trends in Ecology & Evolution 33: 313–325. https://doi.org/10.1016/j.tree.2018.03.002
- Pescador DS, Chacon-Labella J, de la Cruz M, Escudero A (2014) Maintaining distances with the engineer: patterns of coexistence in plant communities beyond the patch-bare dichotomy. New Phytologist 204: 140–148. https://doi.org/10.1111/nph.12899
- Pierce NA, Archer SR, Bestelmeyer BT, James DK (2018) Grass-shrub competition in arid lands: an overlooked driver in grassland-shrubland state transition? Ecosystems. https://doi.org/10.1007/s10021-018-0290-9
- Poulos JM, Rayburn AP, Schupp EW (2014) Simultaneous, independent, and additive effects of shrub facilitation and understory competition on the survival of a native forb (Penstemon palmeri). Plant Ecology: 417–426. https://doi.org/10.1007/s11258-014-0312-4
- Pucheta E, Garcia-Munro VJ, Rolhauser AG, Quevedo-Robledo L (2011) Invasive potential of the winter grass Schismus barbatus during the winter season of a predominantly summer-rainfall desert in Central-Northern Monte. Journal of Arid Environments 75: 390–393. https://doi.org/10.1016/j.jaridenv.2010.11.010
- R Development Core Team (2018) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria.
- Ramírez L, Rada F, Llambí LD (2015) Linking patterns and processes through ecosystem engineering: effects of shrubs on microhabitat and water status of associated plants in the high tropical Andes. Plant Ecology 216: 213–225. https://doi.org/10.1007/s11258-014-0429-5
- Reisner MD, Doescher PS, Pyke DA (2015) Stress-gradient hypothesis explains susceptibility to Bromus tectorum invasion and community stability in North America’s semi-arid Artemisia tridentata wyomingensis ecosystems. Journal of Vegetation Science 26: 1212–1224. https://doi.org/10.1111/jvs.12327
- Rowe HI (2010) Tricks of the trade: techniques and opinions from 38 experts in tallgrass prairie restoration. Restoration Ecology 18: 253–262. https://doi.org/10.1111/j.1526-100X.2010.00663.x
- Salo LF (2005) Red brome (Bromus rubens subsp. madritensis) in North America: possible modes for early introductions, subsequent spread. Biological Invasions 7: 165–180. https://doi.org/10.1007/s10530-004-8979-4
- Schutzenhofer MR, Valone TJ (2006) Positive and negative effects of Erodium cicutarium on an arid ecosystem. Biological Conservation 132: 376–381. https://doi.org/10.1016/j.biocon.2006.04.031
- Seabloom EW, Harpole WS, Reichman OJ, Tilman D (2003) Invasion, competitive dominance, and resource use by exotic and native California grassland species. Proceedings of the Royal Academy of Science 100: 13384–13389. https://doi.org/10.1073/pnas.1835728100
- Sheley RL, James JJ (2014) Simultaneous intraspecific facilitation and interspecific competition between native and annual grasses. Journal of Arid Environments 104: 80–87. https://doi.org/10.1016/j.jaridenv.2014.01.019
- Simberloff D (2006) Invasional meltdown 6 years later: important phenomenon, unfortunate metaphor, or both? Ecology Letters 9: 912–919. https://doi.org/10.1111/j.1461-0248.2006.00939.x
- Simberloff D (2011) Non-natives: 141 scientists object. Nature 475: 35–36. https://doi.org/10.1038/475036a
- Simberloff D, Martin JL, Genovesi P, Maris V, Wardle WA, Aronson J, Courtchamp F, Galil B, Garcia-Bethou E, Pascal M, Pysek P, Sousa R, Tabacchi E, Vila M (2013) Impacts of biological invasions: what’s what and the way forward. Trends in Ecology and Evolution 28: 58–66. https://doi.org/10.1016/j.tree.2012.07.013
- Simberloff D, Von Holle B (1999) Positive interactions of nonindigenous species: invasional meltdown? Biological Invasions 1: 21–32. https://doi.org/10.1023/A:1010086329619
- Simpson A, Eyler MC (2018) First comprehensive list of non-native species established in three major regions of the United States. U.S. Geological Survey Open-File Report 2018–1156. https://doi.org/10.3133/ofr20181156
- Smith DA, Ralls K, Cypher BL, Maldonado JE (2005) Assessment of scat-detection dog surveys to determine kit fox distribution. Wildlife Society Bulletin 33: 897–904. https://doi.org/10.2193/0091-7648(2005)33[897:AOSDST]2.0.CO;2
- St. Clair SB, O’Conner R, Gill R, McMillan B (2016) Biotic resistance and disturbance: rodent consumers regulate post-fire plant invasions and increase plant community diversity. Ecology 97: 1700–1711. https://doi.org/10.1002/ecy.1391
- Stohlgren TJ, Jarnevich C, Chong GW, Evangelista PH (2006) Scale and plant invasions: a theory of biotic acceptance. Preslia 78: 405–426.
- Tewksbury JJ, Lloyd JD (2001) Positive interactions under nurse-plants: spatial scale, stress gradients and foundation species size. Oecologia 127: 425–434. https://doi.org/10.1007/s004420000614
- U.S. Department of the Interior Bureu of Land Management (2011) Check list of plants Carrizo Plain, San Luis Obispo County, California. http://www.blm.gov/ca/st/en/fo/bakersfield/ Programs/carrizo/plants.html [retrieved 11-12-2018]
- Venable DL (2007) Bet hedging in a guild of desert annuals. Ecology 88: 1086–1090. https://doi.org/10.1890/06-1495
- Vila M, Espinar JL, Hejda M, Hulme P, Jarosik V, Maron J, Pergl J, Schaffner U, Sun Y, Pysek P (2011) Ecological impacts of invasive alien plants: a meta-analysis of their effects on species, communities, and ecosystems. Ecology Letters 14: 702–708. https://doi.org/10.1111/j.1461-0248.2011.01628.x
- Vila M, Weiner J (2004) Are invasive plant species better competitors than native plant species? – evidence from pairwise experiments. Oikos 105: 229–238. https://doi.org/10.1111/j.0030-1299.2004.12682.x
- Villareal-Barajas T, Martorell C (2009) Species-specific disturbance tolerance, competition and positive interactions along an anthropogenic disturbance gradient. Journal of Vegetation Science 20: 1027–1040. https://doi.org/10.1111/j.1654-1103.2009.01101.x
- Vitousek PM, D’Antonio CM, Loope LL, Westbrooks R (2017) Biological invasions as global environmental change.
- Webb N, Marshall N, Stringer L, Reed M, Chappell A, Herrick J (2017) Land degradation and climate change: building climate resilience in agriculture. Frontiers in Ecology and the Environment 15: 450–459. https://doi.org/10.1002/fee.1530
- Westphal MF, Noble T, Butterfield HS, Lortie CJ (2018) A test of desert shrub facilitation via radiotelemetric monitoring of a diurnal lizard. Ecology and Evolution 8: 12153–12162. https://doi.org/10.1002/ece3.4673
- Williams DF, Cypher EA, Kelly PA, Miller KJ, Norvell N, Phillips SF, Johnson CD, Colliver GW (1998) Recovery plan for upland species of the San Joaquin Valley, California. Endangered Species Recovery Program 1: 311–319.
- Wright A, Schnitzer SA, Reich PB (2014) Living close to your neighbors: the importance of both competition and facilitation in plant communities. Ecology 95: 2213–2223. https://doi.org/10.1890/13-1855.1
- Zarnetske PL, Gouhier TC, Hacker SD, Seabloom EW, Bokil VA (2013) Indirect effects and facilitation among native and non-native species promote invasion success along an environmental stress gradient. Journal of Ecology 101: 905–915. https://doi.org/10.1111/1365-2745.12093
Supplementary material
Supplementary materials
Data type: multimedia
Explanation note: Supplementary materials for this article are included in Appendix A1, which consists of one supplementary figure (Fig. A1) and seven supplementary tables (Tables A1–A7).