|
Corresponding author: Lubabalo Mofu ( l.mofu@saiab.ac.za ) Academic editor: Emili García-Berthou
© 2019 Lubabalo Mofu, Ross N. Cuthbert, Tatenda Dalu, Darragh J. Woodford, Ryan J. Wasserman, Jaimie T. A. Dick, Olaf L. F. Weyl.
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Mofu L, Cuthbert RN, Dalu T, Woodford DJ, Wasserman RJ, Dick JTA, Weyl OLF (2019) Impacts of non-native fishes under a seasonal temperature gradient are forecasted using functional responses and abundances. NeoBiota 49: 57-75. https://doi.org/10.3897/neobiota.49.34986
|
Abstract
Developing predictive methods to forecast the impacts of existing and emerging invasive species is of critical importance to biodiversity conservation. However, invader impacts are context-dependent, making reliable and robust predictions challenging. In particular, it is unclear how temporal variabilities in relation to temperature regime shifts influence invader ecological impacts. In the present study, we quantify the functional responses of three coexisting freshwater fishes: the native freshwater River Goby Glossogobius callidus, and the non-native Mozambique Tilapia Oreochromis mossambicus and Western Mosquitofish Gambusia affinis, under two temperature treatments using chironomid larvae as prey. This was used along with fish abundance data to determine temporal differences in ecological impacts of each fish species between seasons (i.e. at two corresponding temperatures). All three fish species exhibited potentially population-destabilizing Type II functional responses. Their maximum feeding rates were consistently higher in the warm temperature treatment, whereas attack rates tended to be reduced. Non-native Mozambique Tilapia had the highest maximum feeding rate under both temperature treatments (18 °C and 25 °C), followed by the non-native Western Mosquitofish and lastly the native River Goby, suggesting greater per capita impacts on native prey by non-native fishes. The predatory fish abundances differed significantly according to season, with native River Goby and non-native Mozambique Tilapia generally more abundant than non-native Western Mosquitofish. By multiplying functional response maximum feeding rates with abundances of each fish species across the seasonal gradient, the relative impact potential of non-native Mozambique Tilapia was consistently higher compared to that of native gobies. Western Mosquitofish impacts were less apparent, owing to their low abundances. We demonstrate how seasonal temperature fluctuations affect the relative impact capacities of introduced species and the utility of consumer functional response and the relative impact potential metric in impact forecasting.
Keywords
Context-dependence, impact assessment, introduced species, relative impact potential, seasonal abundance, thermal regime
Introduction
Biological invasions are a central driver of global biodiversity loss (
Despite the considerable work conducted on invasive species, predicting ecological impacts of biological invasions has remained elusive (
Methodological developments, which incorporate native/non-native species resource utilization across context-dependencies, have recently provided robust predictive tools for invasion science (
Classically, the functional response has been combined with the ‘numerical response’ to determine the ‘total response’ of consumers (
The current study focuses on one native and two non-native fish species that co-occur in irrigation ponds within the Sundays River Valley, Eastern Cape, South Africa. These are the native River Goby Glossogobius callidus (Smith, 1937), and two non-native species, Mozambique Tilapia Oreochromis mossambicus (Peters, 1852) and Western Mosquitofish Gambusia affinis (Baird and Girard, 1853). The native River Goby is naturally found in estuarine and freshwater habitats (
Materials and methods
Ethical clearance and permits
The collection of animals and all experiments were carried out in compliance with the Eastern Cape Department of Economic Development and Environmental Affairs (DEDEA permit no. CRO 35/17CR and CRO 36/17CR) and ethical clearance was approved by the National Research Foundation – South African Institute for Aquatic Biodiversity (NRF-SAIAB reference no. 25/4/1/5_2017/03).
Functional response experimental design
River Goby, Mozambique Tilapia and Western Mosquitofish individuals were sourced using a 30 m × 2 m seine net with 12 mm mesh wings and an 8 mm mesh cod-end from Dunbrody (33°27'53"S; 25°33'02"E) and Disco Chicks (33°27'26"S; 25°39'57"E) irrigation ponds, Eastern Cape, South Africa. Upon capture, fish were transported to NRF-SAIAB, Grahamstown in continuously aerated containers with source water. Each fish species was housed separately in a controlled temperature and light laboratory and kept under a 12:12 light:dark cycle. Temperature was maintained at either 18 ± 2 °C or 25 ± 2 °C (i.e. experimental temperature groups) for seven days prior to experimentation, with each species acclimated separately in 40 L fish tanks in a closed recirculating system. All fish were maintained on a standardized diet of larval chironomids ad libitum. The chironomid larvae were collected by kick sampling from the Bloukrans River (33°19'06"S; 26°34'22"E) using a kick net (1000 µm). The chironomids were then strained twice through 2.0 mm and then 1.0 mm sieves to obtain the experimental size class (total length (TL) ± standard deviation ((SD) 1.5 ± 0.11 mm) and then rinsed thoroughly with deionized water to remove any other food sources.
Functional response experiments were performed at 18 °C and 25 °C, reflecting respective spring and summer temperatures at the sampling locations. Following
Fish abundances
The fish predator abundance data were obtained from the NRF-SAIAB`s monitoring program of irrigation ponds in the Sundays River Valley, Eastern Cape, South Africa. Abundance from two irrigation ponds were used, ML Swart (33°24'33"S; 25°29'04"E), and River Bend (33°26'23"S; 25°42'25"E). The pond names represent either the property or farm owner’s name, as recorded by the Lower Sundays River Water User Association. These ponds were selected on the basis that they were surveyed in both spring and summer and that all three species were captured to give abundance estimates. During each survey, the irrigation pond water temperatures were measured using a HANNA HI98129 combo pH and electrical conductivity meter (HANNA Instruments Inc., Woonsocket, USA). Spring (18 °C) and summer (25 °C) abundance estimates were used in this study as they were in line with the experimental temperatures, and reflect seasonal temperature means.
The ponds were surveyed using a 30 m × 2 m siene net with 12 mm mesh wings and an 8 mm mesh cod-end. At least three hauls were conducted per pond and, upon completion of a single haul, all fish were kept alive in a continuously aerated container (20 L) until every seine haul was completed within a pond. Fish were then identified to species-level, enumerated and released back to the water. The abundance data were based on maximum catch field abundances using mean catch per 100 m2.
Statistical analyses
Generalized linear models (GLMs) assuming a Poisson error distribution and log link were used to analyze overall prey consumption with respect to species, temperature and prey supply. Likewise, GLMs were used to compare fish abundances with respect to species, season and pond. Non-significant terms and interactions were removed stepwise to obtain minimum adequate models (
To distinguish between Type II and III functional responses, logistic regression of the proportion of prey consumed as a function of initial prey density was performed (but see also
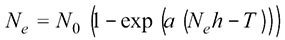 (1)
(1)
where Ne is the number of prey eaten, N0 is the initial density of prey, a is the attack rate, h is the handling time and T is the experiment duration (fixed at 1). To enable model fitting, the Lambert W function was used (needed as Ne appears on both sides of the equation; (
We calculated relative impact potential (RIP) of native (i.e. River Goby) and non-native (i.e. Mozambique Tilapia, Western Mosquitofish) species using the mean bootstrapped functional response maximum feeding rate (FR) and abundance (AB) for the three species at each season and pond (
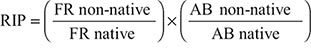 (2)
(2)
when RIP < 1, the predicted impact of the non-native fish is predicted to be less than the native; when RIP = 1, there is no difference in impact between the fish species; whereas when RIP > 1, the non-native has a greater impact than the native. To integrate uncertainty into the RIP score, a probability density function (pdf) was applied using the standard deviation (SD) of the FR and AB estimates and this generated 80% confidence intervals (CIs) (see
Results
Functional response
Prey survival of larval chironomids was 99% in control groups with predators absent, and thus prey mortality in the experimental groups was attributed to predation. Overall consumption was significantly different among fish species (χ2 = 221.67, df = 2, p < 0.001). Native River Goby consumed significantly fewer prey than both non-native Mozambique Tilapia (z = 14.61, p < 0.001) and non-native Western Mosquitofish (z = 8.43, p < 0.001). Mozambique Tilapia, in turn, consumed significantly more prey than Western Mosquitofish overall (z = 6.41, p < 0.001). Consumption was also significantly greater at the higher temperature, analogous with the summer season (χ2 = 179.61, df = 1, p < 0.001), and consumption increased with temperature for all species as there was no significant ‘predator × temperature’ interaction (χ2 = 3.54, df = 2, p = 0.171; Figure
At 18 °C (i.e. spring temperature), all three fish species displayed a Type II functional response (Table
At 25 °C (i.e. summer temperature), all three fish species also exhibited a Type II functional response (Table

Functional response curves for native River Goby (blue circles, solid lines), non-native Mozambique Tilapia (red squares, dashed lines) and non-native Western Mosquitofish (green triangles, dotted lines) at 18 °C (a) and 25 °C (b). Means are ± SE. Filled points are means and unfilled points are raw data.
Parameter estimates from first-order logistic regression of the proportion of consumed prey as a function of prey density, with rounded functional response estimates, a = attack rate; h = handling time, 1/h = maximum feeding rate.
| Predator | Temperature | First-order term, p | a | p | h | p | 1/h |
| Native River Goby | 18 °C | –0.04, <0.001 | 4.34 | <0.001 | 0.05 | <0.001 | 20.00 |
| Non-native Mozambique Tilapia | 18 °C | –0.03, <0.001 | 5.23 | <0.001 | 0.02 | <0.001 | 43.48 |
| Non-native Western Mosquitofish | 18 °C | –0.03, <0.001 | 4.74 | <0.001 | 0.04 | <0.001 | 27.78 |
| Native River Goby | 25 °C | –0.03, <0.001 | 3.65 | <0.001 | 0.03 | <0.001 | 34.48 |
| Non-native Mozambique Tilapia | 25 °C | –0.01, <0.001 | 2.20 | <0.001 | 0.01 | <0.001 | 111.11 |
| Non-native Western Mosquitofish | 25 °C | –0.02, <0.001 | 3.80 | <0.001 | 0.02 | <0.001 | 58.82 |
Fish abundances
There was a significant ‘species × season × pond’ interaction (χ2 = 92.54, df = 2, p < 0.001; Figure

Relative impact potential
Under both spring and summer treatments, the non-native Mozambique Tilapia consistently displayed relative impact potential scores > 1 relative to the native River Goby irrespective of focal ponds, suggesting greater impact than the native species (Table
Relative Impact Potential (RIP) using mean bootstrapped maximum feeding rates for non-native Mozambique Tilapia and non-native Western Mosquitofish against native River Goby. Field abundance data are integrated from ML Swart and River Bend ponds in spring and summer. Uncertainties are reflected through 80% confidence intervals (CIs).
| Species | Season | Pond | Mean FR maximum feeding ± SD | Mean field abundance ± SD | RIP | CIs | P rip>1 |
| Non-native Mozambique Tilapia, native River Goby | Spring | ML Swart | 45.40 ± 11.31, 19.96 ± 3.53 | 2.41 ± 2.84, 2.25 ± 3.10 | 7.25 | 0.42 – 16.32 | 75.17 |
| Non-native Mozambique Tilapia, native River Goby | Spring | River Bend | 45.40 ± 11.31, 19.96 ± 3.53 | 2.04 ± 0.69, 5.22± 4.80 | 1.69 | 0.35 – 3.57 | 55.21 |
| Non-native Western Mosquitofish, native River Goby | Spring | ML Swart | 26.68 ± 2.87, 19.96 ± 3.53 | 0.19 ± 0.17, 2.25 ± 3.10 | 0.35 | 0.29 – 0.78 | 70.78 |
| Non-native Western Mosquitofish, native River Goby | Spring | River Bend | 26.68 ± 2.87, 19.96 ± 3.53 | 2.04 ± 0.69, 5.22 ± 4.80 | 1.01 | 0.22 – 2.11 | 33.40 |
| Non-native Mozambique Tilapia, native River Goby | Summer | ML Swart | 125.02 ± 54.57, 32.60 ± 4.10 | 15.50 ± 4.80, 13.70 ± 11.10 | 7.30 | 1.58 – 15.35 | 96.40 |
| Non-native Mozambique Tilapia, native River Goby | Summer | River Bend | 125.02 ± 54.57, 32.60 ± 4.10 | 20.10 ± 8.02, 5.41 ± 2.77 | 18.26 | 5.21 – 36.15 | 99.98 |
| Non-native Western Mosquitofish, native River Goby | Summer | ML Swart | 97.17 ± 148.60, 32.60 ± 4.10 | 20.70 ± 21.20, 13.70 ± 11.10 | 7.58 | 0.30 – 16.56 | 69.64 |
| Non-native Western Mosquitofish, native River Goby | Summer | River Bend | 97.17 ± 148.60, 32.60 ± 4.10 | 0.06 ± 0.13, 5.41 ± 2.77 | 0.05 | 0.00 – 0.09 | 40.20 |
The relative impact potential biplots concur with the relative impact potential scores (Figure

Relative impact potential (RIP) biplots (see also Table
Discussion
Using the relative impact potential metric proposed by
Temperature differences had a significant effect on the functional response parameters, wherein attack rates were high in spring (i.e. 18 °C) and were reduced in summer (i.e. 25 °C). This result concurs with
Secondly, we show that there was significant variation in fish abundances among species according to season, and also between ponds. Such variation in fish abundances seems to be a common theme, especially in fish communities that co-occur in environments and this is driven by spatial and temporal variation in life-history traits (
Changes in relative impact potential scores with seasonal temperature fluctuations and fish abundances from different localities demonstrate how such context-dependencies can have a critical effect on the relative field impact capacities of introduced species through time (
The present study further demonstrates the usefulness of numerical response proxies such as abundances in rapid assessments of potential impacts of introduced species. Indeed, in many cases, impact predictions are inherently limited if based on per capita impacts alone, given the importance of abundances in discernments of overall offtake rates by consumer populations (
Overall, this study provides further evidence of the strength of the relative impact potential metric in predicting ecological impacts of species and provides an extension to the framework by integrating an environmental gradient, which reflects seasonal temperature fluctuations. The identification of temporal shifts in impact across seasons and habitats in our study presents novel insights into invader impact. In many ecosystems, data on species abundances are still lacking, but since the relative impact potential metric enables impact predictions for species without invasion histories, we recommend more surveys to estimate abundance of potential invaders and/or for practitioners to incorporate other proxies (such as fecundity) into the metric (see
Acknowledgements
This study forms part of a PhD research project supported by the Professional Development Programme Doctoral Scholarship with the South African Institute for Aquatic Biodiversity (NRF, Grant No. 101039), the CIB/DST Centre of Excellence for Invasion Biology (CIB) and the National Research Foundation – South African Research Chairs Initiative of the Department of Science and Technology (Inland Fisheries and Freshwater Ecology, Grant No. 110507) and NRF incentive funding (Grant Nos. 109015 to O.L.F.W., 103581 to D.J.W and 88746 to R.J.W). We acknowledge use of infrastructure and equipment provided by the NRF-SAIAB Research Platform and the funding channelled through the NRF-SAIAB Institutional Support system. Eastern Cape Department of Economic Development and Environmental Affairs (DEDEA) is thanked for issuing research permits. Any opinion, finding and conclusion or recommendation expressed in this material is that of the authors. Consequently, the National Research Foundation of South Africa and CapeNature do not accept any liability in this regard.
References
- Adams PA (1980) Consumer functional response and competition in consumer-resource systems. Theoretical Population Biology 17: 80–102. https://doi.org/10.1016/0040-5809(80)90016-7
- Alexander ME, Dick JTA, Weyl OLF, Robinson TB, Richardson DM (2014) Existing and emerging high impact invasive species are characterized by higher functional responses than natives. Biological Letters 10: 2–6. https://doi.org/10.1098/rsbl.2013.0946
- Amezcua F, Amezcua-Linares F (2014) Seasonal changes of fish assemblages in a subtropical lagoons in the SE Gulf of California. The Scientific World Journal 15: 1–14. https://doi.org/10.1155/2014/968902
- Barrios-O`Neill D, Kelly R, Dick JTA, Ricciardi A, MacIsaac HJ, Emmerson MC (2016) On the context-dependent scaling of consumer feeding rates. Ecology Letters 19: 668–678. https://doi.org/10.1111/ele.12605
- Bertelsmeier C, Keller L (2018) Bridgehead effects and role of adaptive evolution in invasive populations. Trends in Ecology & Evolution 33: 527–534. https://doi.org/10.1016/j.tree.2018.04.014
- Bolker BM (2008) Ecological models and data in R. Priceton University Press, Princeton, New Jersey. https://doi.org/10.2307/j.ctvcm4g37
- Canonico GC, Arthington A, Mccrary JK, Thieme ML (2005) The effects of introduced tilapias on native biodiversity. Aquatic Conservation: Marine and Freshwater Ecosystems 15: 463–483. https://doi.org/10.1002/aqc.699
- Chapin III FS, Zavaleta ES, Eviner VT, Naylor RL, Vitousek PM, Reynolds HL, Hooper DU, Lavorel S, Sala OE, Hobbie SE, Mack MC, Díaz S (2000) Consequences of changing biodiversity. Nature 405: 234–242. https://doi.org/10.1038/35012241
- Clarke A, Johnston NM (1999) Scaling of metabolic rate with body mass and temperature in teleost fish. Journal of Animal Ecology 68: 893–905. https://doi.org/10.1046/j.1365-2656.1999.00337.x
- Crawley MJ (2007) The R Book. John Wiley and Sons, Chichester, UK.
- Cuthbert RN, Dick JTA, Callaghan A, Dickey JWE (2018) Biological control agent selection under environmental change using functional responses, abundances and fecundities; the Relative Control Potential (RCP) metric. Biological Control 121: 50–57. https://doi.org/10.1016/j.biocontrol.2018.02.008
- Cuthbert RN, Dickey JWE, Coughlan NE, Joyce PWS, Dick JTA (in press) The functional response ratio (FRR): advancing comparitive metrics for predicting the ecological impacts of invasive alien species. Biological Invasions.
- Dick JTA, Alexander ME, Jeschke JM, Ricciardi A, MacIssac HJ, Robinson TB, Kumschick S, Weyl OLF, Dunn AM, Hatcher MJ, Paterson RA, Farnsworth KD, Richardson DM (2014) Advancing impact prediction and hypothesis testing in invasion ecology using a comparative functional response approach. Biological Invasions 16: 735–753. https://doi.org/10.1007/s10530-013-0550-8
- Dick JTA, Gallagher K, Avlijas S, Clarke HC, Lewis SE, Leung S, Minchin D, Caffrey J, Alexander ME, Maguire C, Harrod C, Reid N, Haddaway NR, Farnsworth KD, Penk M, Ricciardi A (2013) Ecological impacts of an invasive predator explained and predicted by comparative functional responses. Biological Invasions 15: 837–846. https://doi.org/10.1007/s10530-012-0332-8
- Dick JTA, Laverty C, Lennon JJ, Barrios-O’Neill D, Mensik PJ, Britton JR, Médoc V, Boets P, Alexander ME, Taylor NG, Dunn AM, Hatcher MJ, Rosewarne PJ, Crookes S, MacIsaac HJ, Xu M, Ricciardi A, Wasserman RJ, Ellender BR, Weyl OLF, Lucy FE, Banks PB, Dodd JA, MacNeil C, Penk MR, Aldridge DC, Caffrey JM (2017) Invader Relative Impact Potential: a new metric to understand and predict the ecological impacts of existing, emerging and future invasive alien species. Journal of Applied Ecology 54: 1259–1267. https://doi.org/10.1111/1365-2664.12849
- Dickey JWE, Cuthbert RN, Rea M, Laverty C, Crane K, South J, Briski E, Chang X, Coughlan NE, MacIsaac HJ, Ricciardi A, Riddell GE, Xu M, Dick JTA (2018) Assessing the relative potential ecological impacts and invasion risk of emerging and future invasive alien species. Neobiota 40: 1–24. https://doi.org/10.3897/neobiota.40.28519
- Engelbrecht GD, Mulder PFS (1999) Extremely high genetic differentiation between two populations of the River Goby, Glossogobius callidus (Smith, 1937). Water SA 25: 85–90.
- Englund G, Öhlund G, Hein CL, Diehl S (2011) Temperature dependence of the functional response. Ecology Letters 14: 914–921. https://doi.org/10.1111/j.1461-0248.2011.01661.x
- Fussmann KE, Schwarzmüller F, Brose U, Jousset A, Rall BC (2014) Ecological stability in response to warming. Nature Climate Change 4: 206–210. https://doi.org/10.1038/nclimate2134
- Gallardo B, Clavero M, Sánchez MI, Vilá M (2016) Global ecological impacts of invasive species in aquatic ecosystems. Global Change Biology 22: 151–163. https://doi.org/10.1111/gcb.13004
- Grigaltchik VS, Ward AJW, Seebacher F (2012) Thermal acclimation of interactions: differential responses to temperature change alter predator-prey relationship. Proceedings of the Royal Society 279: 4058–4064. https://doi.org/10.1098/rspb.2012.1277
- Havel JE, Kovalenko KE, Thomaz SM, Amalfitano S, Kats LB (2015) Aquatic invasive species: challenges for the future. Hydrobiologia 750: 147–170. https://doi.org/10.1007/s10750-014-2166-0
- Holling CS (1959) Some characteristics of simple types of predation and parasitism. Canadian Entomologist 91: 385–398. https://doi.org/10.4039/Ent91385-7
- Hothorn T, Bretz F, Westfall P (2008) Simultaneous inference in general parametric models. Biometrical Journal 50: 346–363. https://doi.org/10.1002/bimj.200810425
- Iacarella JC, Dick JTA, Alexander MW, Ricciardi A (2015) Ecological impacts of invasive species along temperature gradients: testing the role of environmetal matching. Ecological Application 25: 706–716. https://doi.org/10.1890/14-0545.1
- IUCN (2006) IUCN Red list of threatened species.
- James NC, Cowley PD, Whitfield AK, Lamberth SJ (2007) Fish communities in temporal open/closed estuaries from the warm- and cool-temperate regions of South Africa: a review. Reviews in Fish and Biology and Fisheries 17: 565–580. https://doi.org/10.1007/s11160-007-9057-7
- Jeschke JM, Kopp M, Tollrian R (2004) Consumer-food systems: why type I functional responses are exclusive to filter feeders. Biological Reviews 89: 337–349. https://doi.org/10.1017/S1464793103006286
- Jobling M (1981) Temperature tolerance and the final preferendum-rapid methods for the assessment of optimum growth temperatures. Journal of Fish Biology 19: 439–455. https://doi.org/10.1111/j.1095-8649.1981.tb05847.x
- Juliano SA (2001) Nonlinear curve fitting: predation and functional response curves. In: Scheiner SM, Gurevitch J (Eds) Design and analysis of ecological experiments Oxford University Press. 178–196.
- Kalinkat G, Schneider FD, Digel C, Guill C, Rall BC, Brose U (2013) Body masses, functional responses and predator-prey stability. Ecology Letters 16: 1126–1134. https://doi.org/10.1111/ele.12147
- Lang B, Ehnes RB, Brose U, Björn CR (2017) Temperature and consumer type dependencies of energy flows in natural communities. Oikos 126: 1717–1725. https://doi.org/10.1111/oik.04419
- Latombe G, Pyšek P, Jeschke JM, Blackburn TM, Bacher S, Capinha C, Costello MJ, Fernández M, Gregory RD, Hobern D, Hui C, Jetz W, Kumschick S, McGrannachan C, Pergl J, Roy HE, Scalera R, Squires ZE, Wilson JRU, Winter M, Genovesi P, McGeoch MA (2017) A vision for global monitoring of biological invasions. Biological Conservation 213: 295–308. https://doi.org/10.1016/j.biocon.2016.06.013
- Laverty C, Dick JTA, Alexander ME, Lucy FE (2015) Differential ecological impacts of invader and native predatory freshwater amphipods under environmental change are revealed by comparative functional responses. Biological Invasions 17: 1761–1770. https://doi.org/10.1007/s10530-014-0832-9
- Laverty C, Green KD, Dick JTA, Barrios-O’Neill D, Mensik PJ, Médoc V, Spataro T, Caffrey JM, Lucy FE, Boets P, Britton JR, Pegg J, Gallagher C (2017) Assessing the ecological impacts of invasive species based on their functional responses and abundances. Biological Invasions 19: 1653–1665. https://doi.org/10.1007/s10530-017-1378-4
- Maddern MG, Morgan DL, Gill HS (2007) Distribution, diet and potential ecological impacts of the introduced Mozambique mouthbrooder Oreochromis mossambicus Peters (Pisces: Cichlidae) in Western Australia. Journal of the Royal Society of Western Australia 90: 203–214.
- Mofu L, Woodford DJ, Wasserman RJ, Tatenda D, Weyl OLF (in press) Diet of Glossogobius callidus (Teleostei: Gobiidae) in freshwater impoundments in the Sundays River Valley of the Eastern Cape, South Africa. African Journal of Aquatic Sciences.
- Murdoch WW, Oaten A (1975) Predation and population stability. Advances in Ecological Research 9: 1–131. https://doi.org/10.1016/S0065-2504(08)60288-3
- O’Gorman EJ, Zhao L, Pichler DE, Adams G, Friberg N, Rall BC, Seeney A, Zhang H, Reuman DC, Woodward G (2017) Unexepected changes in community size structure in a natural warming experiment. Nature Climate Change 7: 659–663. https://doi.org/10.1038/nclimate3368
- Paine RT (1980) Food webs: linkage, interaction strength and community infrastructure. Journal of Animal Ecology 49: 667–685. https://doi.org/10.2307/4220
- Penk M, Saul WC, Dick JTA, Donohue I, Alexander ME, Linzmaier S, Jeschke JM (2017) A trophic interaction framework for identifying the invasive capacity of novel organisms. Methods in Ecology and Evolution 8: 1786–1794. https://doi.org/10.1111/2041-210X.12817
- Pritchard DW, Paterson RA, Bovy HC, Barrios-O’Neill D (2017) FRAIR : an R package for fitting and comparing consumer functional responses. Methods in Ecology and Evolution 8: 1528–1534. https://doi.org/10.1111/2041-210X.12784
- Pyke GH (2008) Plague minnow or mosquito Fish ? A review of the biology and impacts of introduced Gambusia species. Annual Review of Ecology, Evolution, and Systematics 39: 171–191. https://doi.org/10.1146/annurev.ecolsys.39.110707.173451
- Rall BC, Brose U, Hartvig M, Kalinkat G, Schwarzmüller F, Vucic-Pestic O, Petchey OL (2012) Universal temperature and body-mass scaling of feeding rates. Philosophical Transactions of the Royal Society B 367: 2923–2934. https://doi.org/10.1098/rstb.2012.0242
- Rall BC, Vucic-Pestic O, Ehnes RB, Emmerson M, Brose U (2010) Temperature, predator-prey interaction strength and population stability. Global Change Biology 16: 2145–2157. https://doi.org/10.1111/j.1365-2486.2009.02124.x
- Ricciardi A, Hoopes MF, Marchetti MP, Lockwood JL (2013) Progress toward understanding the ecological impacts of nonnative species. Ecological Monographs 83: 263–282. https://doi.org/10.1890/13-0183.1
- Ricciardi A, Blackburn TM, Carlton JT, Dick JTA, Hulme PE, Iacarella JC, Jeschke JM, Liebhold AM, Lockwood JL, MacIsaac HJ, Pyšek P, Richardson DM, Ruiz GM, Simberloff D, Sutherland WJ, Wardle DA, Aldridge DC (2017) Invasion science: A horizon scan of emerging challenges and opportunities. Trends in Ecology & Evolution 32: 464–474. https://doi.org/10.1016/j.tree.2017.03.007
- Rip JMK, McCann KS (2011) Cross-ecosystem differerences in stability and the principle of energy flux. Ecology Letters 14: 733–740. https://doi.org/10.1111/j.1461-0248.2011.01636.x
- Roessig JM, Woodley CM, Cech Jr JJ, Hansen LJ (2004) Effects of global climate change on marine and estuarine fishes and fisheries. Reviews in Fish Biology and Fisheries 14: 251–275. https://doi.org/10.1007/s11160-004-6749-0
- Rogers A (1972) Random search and insect population models. Journal of Animal Ecology 41: 369–383. https://doi.org/10.2307/3474
- Rosenbaum B, Rall BC (2018) Fitting functional responses: direct parameter estimation by simulating differential equations. Methods in Ecology and Evolution 9: 2076–2090. https://doi.org/10.1111/2041-210X.13039
- Sala O, Chapin III FS, Armesto JJ, Berlow E, Bloomfield J, Dirzo R, Huber-Sanwald E, Huenneke LF, Jackson RB, Kinzig A, Leemans R, Lodge DM, Mooney HA, Oesterheld M, Poff NL, Sykes MT, Walker BH, Walker M, Wall DH (2000) Global biodiversity scenarios for the year 2100. Science 287: 1770–1776. https://doi.org/10.1126/science.287.5459.1770
- Sanford E (1999) Regulation of keystone predation by small changes in ocean temperature. Science 283: 2095–2097. https://doi.org/10.1126/science.283.5410.2095
- Sanford E (2002) The feeding, growth, and eneretics of two rocky intertidal predators (Pisaster ochracues and Nucella canaliculata) under water temperatures simulating episodic upwelling. Journal of Experimental Marine Biology and Ecology 273: 199–218. https://doi.org/10.1016/S0022-0981(02)00164-8
- Sarnelle O, Wilson AE (2008) Type III functional response in Daphnia. Ecology 89: 1723–1732. https://doi.org/10.1890/07-0935.1
- Seebens H, Blackburn TM, Dyer EE, Genovesi P, Hulme PE, Jeschke JM, Pagad S, Pyšek P, van Kleunen M, Winter M, Ansong M, Arianoutsou M, Bacher S, Blasius B, Brockerhoff EG, Brundu G, Caphinha C, Causton CE, Celesti-Grapow L, Dawson W, Dullinger S, Economo EP, Fuentes N, Guénard B, Jäger H, Kartesz J, Kenis M, Kühn I, Lenzner B, Liebhold AM, Mosena A, Moser D, Nentwing W, Nishino M, Pearman D, Pergl J, Rabitsch W, Rojas-Sandoval J, Roques A, Rorke S, Rossinelli S, Roy HE, Scalera R, Schindler S, Štajerova K, Tokarska-Guzik B, Walker K, Ward DF, Yamanaka T, Essl F (2018) Global rise in emerging alien species results from increased accessibility of new source pools. Proceedings of the National Academy of Science 155: E2264–E2273. https://doi.org/10.1073/pnas.1719429115
- Sentis A, Hemptinne JL, Brodeur J (2012) Using functional response modelling to investigate the effect of temperature on predator feeding rate and energetic effieciency. Oecologia 169: 1117–1125. https://doi.org/10.1007/s00442-012-2255-6
- Shuai F, Lek S, Li X, Zhao T (2018) Biological invasions undermine the functional diversity of fish community in large subtropical river. Biological Invasions 20: 2981–2996. https://doi.org/10.1007/s10530-018-1751-y
- Simberloff D, Martin J-L, Genovesi P, Maris V, Mardle DA, Aronson J, Courchamp F, Galil B, García-Berthou E, Pascal M, Pyšek P, Sousa R, Tabacchi E, Vilà M (2013) Impacts of biological invasions: what’s what and the way forward. Trends in Ecology & Evolution 28: 58–66. https://doi.org/10.1016/j.tree.2012.07.013
- Skelton PH (2001) A complete guide to the freshwater fishes of Southern Africa (2nd edn). Struik Publishers, Cape Town, 365 pp.
- Solomon ME (1949) The natural control of animal populations. Journal of Animal Ecology 18: 1–35. https://doi.org/10.2307/1578
- Sorte CJB, Ibáñez I, Blumenthal DM, Molinari NA, Miller LP, Grosholz ED, Diez JM, D`Antonio CM, Olden JD, Jones SJ, Dukes JS (2013) Poised to proser? A cross-system comparison of climate change effects on native and non-native species perfomance. Ecology Letters 16: 261–270. https://doi.org/10.1111/ele.12017
- South J, Dick JTA, McCard M, Barrios-O’Neill D, Anton A (2017) Predicting predatory impact of juvenile invasive lionfish (Pterois volitans) on a crustacean prey using functional response analysis: effects of temperature, habitat complexity and light regimes. Environmental Biology of Fishes 100: 115–1165. https://doi.org/10.1007/s10641-017-0633-y
- Thomsen MS, Olden JD, Wernberg T, Griffin JN, Silliman BR (2011) A broad framework to organize and compare ecological invasion impacts. Environmental Research 111: 899–908. https://doi.org/10.1016/j.envres.2011.05.024
- Turak E, Harrison I, Dudgeon D, Abell R, Bush A, Darwall W, Finlayson CM, Ferrier S, Freyhof J, Hermoso V, Juffe-Bignoli D, Linke S, Nel J, Patricio HC, Pittock J, Raghavan R, Revenga C, Simaika JP, De Wever A (2016) Essential biodiversity variables for measuring change in global freshwater biodiversity. Biological Conservation 213: 272–279. https://doi.org/10.1016/j.biocon.2016.09.005
- Uiterwaal SF, DeLong JP (2018) Multiple factors, including arean size, shape the functional responses of ladybird beetles. Journal of Applied Ecology 55: 2429–2438. https://doi.org/10.1111/1365-2664.13159
- Uszko W, Diehl S, Englund G, Amarasekare P (2017) Effects of warming on predator-prey interactions – a resource based approach and a theoretical synthesis. Ecology Letters 20: 513–523. https://doi.org/10.1111/ele.12755
- Viherluoto M, Viitasalo M (2001) Effects of light on the feeding rates of pelagic and littoral mysid shrimps: a trade-off between feeding success and predation avoidance. Journal of Experimental Marine Biology and Ecology 261: 237–256. https://doi.org/10.1016/S0022-0981(01)00277-5
- Vitousek PM, D`Antonio CM, Loope LL, Westbrooks R (1996) Biological invasions as global environmental change. American Naturalist 84: 468–478.
- Vucic-Pestic O, Birkhofer K, Rall BC, Scheu S, Brose U (2010a) Habitat structure and prey aggregation determine the functional response in a soil predator-prey interaction. Pedobiologia 53: 307–312. https://doi.org/10.1016/j.pedobi.2010.02.003
- Vucic-Pestic O, Rall BC, Kalinkat G, Brose U (2010b) Allometric functional response model: body masses constrain interaction strengths. Journal of Animal Ecology 79: 249–256. https://doi.org/10.1111/j.1365-2656.2009.01622.x
- Wasserman RJ (2012) Feeding ecology of the early life-history stages of two dominant gobiid species in the headwater of a warm-temperature eastuary. Easuarine, Coastal and Shelf Science 109: 11–19. https://doi.org/10.1016/j.ecss.2012.05.003
- Wasserman RJ, Alexander ME, Weyl OL, Barrios‐O’Neill D, Froneman PW, Dalu T (2016) Emergent effects of structural complexity and temperature on predator-prey interactions. Ecosphere 7: e01239. https://doi.org/10.1002/ecs2.1239
- Woodford DJ, Hui C, Richardson DM, Weyl OLF (2013) Propagule pressure drives establishment of introduced freshwater fish: quantitative evidence from an irrigation network. Ecological Applications 23: 1926–1937. https://doi.org/10.1890/12-1262.1
- Zengeya TA, Booth AJ, Bastos ADS, Chimimba CT (2011) Trophic interrelationships between the exotic Nile tilapia, Oreochromis niloticus and indigenous tilapiine cichlids in a subtropical African river system (Limpopo River, South Africa). Environmental Biology of Fishes 92: 479–489. https://doi.org/10.1007/s10641-011-9865-4