|
Corresponding author: Josie South ( josiesouth93@gmail.com ) Academic editor: Anthony Ricciardi
© 2019 Josie South, Monica McCard, Dumisani Khosa, Lubabalo Mofu , Takudzwa C. Madzivanzira, Jaimie T. A. Dick, Olaf L. F. Weyl.
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
South J, McCard M, Khosa D, Mofu L, Madzivanzira TC, Dick JTA, Weyl OLF (2019) The effect of prey identity and substrate type on the functional response of a globally invasive crayfish. NeoBiota 52: 9-24. https://doi.org/10.3897/neobiota.52.39245
|
Abstract
Biological invasions threaten biodiversity on a global scale, therefore, developing predictive methods to understand variation in ecological change conferred is essential. Trophic interaction strength underpins community dynamics, however, these interactions can be profoundly affected by abiotic context, such as substrate type. The red swamp crayfish (Procambarus clarkii) has successfully invaded a number of freshwater ecosystems. We experimentally derive the Functional Response (FR) (density dependent predation) of the red swamp crayfish preying upon both a benthic prey; chironomid larvae, and a pelagic prey; Daphnia magna, on a no substrate control, sand, and gravel substrates to determine whether (1) there is a higher impact on prey that are benthic, and (2) whether the presence of different substrate types can dampen the interaction strength. We apply and demonstrate the utility of the Functional Response Ratio (FRR) metric in unravelling differences in ecological impact not obvious from traditional FR curves. Procambarus clarkii is capable of constantly utilising high numbers of both benthic and pelagic prey items, showing a Type II functional response under all scenarios. The presence of gravel and sand substrate each independently decreased the magnitude FR upon D. magna. Though, with regards to chironomid larvae the FR curves showed no difference in magnitude FR, the FRR reveals that the highest impact is conferred when foraging on sand substrate. This reinforces the need for impact assessments to be contextually relevant.
Keywords
Aquatic invasions, functional response ratio, invader impact, macroinvertebrates, Procambarus clarkii, substrate
Introduction
Aquatic biological invasions are increasing at an alarming rate driven by increased connectivity due to new trade routes and ongoing climatic change (
Invasive species are often characterised by their efficiency of resource consumption, whereby they generally show a higher per capita effect towards a focal resource in comparison to a native analogue (
Species usually form habitat associations depending on their specific life history needs or due to optimal foraging theory, wherein the most productive habitat is selected (
The red swamp crayfish, Procambarus clarkii (Girard, 1852), is a widespread invasive species due to its wide thermal tolerance, high trait plasticity ,and popularity in aquaculture and the pet trade (
Methods
Specimens of sub-adult P. clarkii (mean ± SD: 21.62 ± 2.6 mm carapace length, n = 25) were supplied by Seahorse Aquariums (Republic of Ireland) and maintained at the Queen’s University Marine Laboratory in a continuously aerated 600 L holding tank, with shelters made of drainpipe halves, supplied with dechlorinated tap water held at 23 ± 1.0 °C using aquarium heaters and subject to natural light regime. During the experimental period there was no cannibalism. Procambarus clarkii were maintained on commercial sinking fish food (JBL, Germany) to avoid conditioning to experimental prey items. As this species breeds at small sizes and because they are controlled species within the United Kingdom and Northern Ireland non-reproductive sub-adult crayfish were used in this experiment. Two live prey species, chironomid larvae (3.0 ± 0.6 SD mm total length) (Chironomidae) and Daphnia magna (0.3 ± 0.01 SD mm) (Daphnia magna; Daphniidae), were supplied, also from Seahorse Aquariums. Appropriate ethical approval for the use of these animals in research was obtained from the QUB Ethics Committee (School of Biological Sciences) and National Research Foundation – South African Institute for Aquatic Biodiversity (NRF–SAIAB ethics boards (25/4/1/5_2018-06).
Functional Response Experiment
A full factorial experimental design was employed to test differences in FR with regards to factors: “prey species” (2 levels), “density” (9 levels), and “substrate” (3 levels). Experimental arenas (W: 20 × L: 30 × H: 14 cm, 8 L) were held at 23 ± 1.0 °C and experiments were completed in a fully randomised design. Three substrate treatments were supplied: no substrate, commercial aquarium sand supplied at a depth of 4 mm in the experimental arena, and gravel (grain size: 8 × 4 mm) positioned using one 4 mm layer at the bottom of the experimental arena. Arenas were covered from the top and sides to avoid any synergistic or antagonistic conspecific effects. Individual predators were added per arena and allowed to acclimate for one hour before adding prey items. Each prey species was supplied at 9 densities (2, 7, 15, 40, 60, 90, 120, 200, 300 individuals per arena, n = 5 for each treatment), crayfish were allowed to feed for 1 h, after which the predators were removed, and number of prey items consumed were enumerated.
Control trials were carried out in experimental tanks at identical environmental conditions, wherein each prey species was supplied at the highest density in the absence of predators (n = 3 per substrate treatment) to determine potential background mortality. Each crayfish was re-used multiple times, but experienced each density of each prey type only once. Crayfish were given at least 3 days between use and were not fed for 24 h prior to experimental procedure to standardise hunger levels.
All analyses were undertaken using the R Statistical Software (v. 3.4.3). A generalised linear model (GLM) with a quasi-Poisson error distribution was used to determine differences in prey consumption with regards to factors “prey species”, “prey density”, and “substrate complexity”, using a Type 3 Anova and χ2 to report the effect size of a factor on the dependent variable. Tukey’s HSD was used to generate pairwise estimates with Holm-Bonferroni adjustment of P values post-hoc.
The R package ‘frair’ (
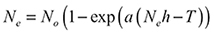 (1)
(1)
wherein, Ne represents the number of prey eaten, N0 is the initial density of prey, a is the attack parameter, h is the handling parameter, and T is the total time available. The Lambert W function was implemented to fit the model to the data (
The FRR (
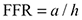 (2)
(2)
where a is the attack parameter and h is the handling parameter derived from the FR curve. In this case the FRR is used as a diagnostic tool to determine whether there were differences that can not be observed from the usual FR outputs. Therefore, the higher the FRR value the higher the inferred impact (
Results
Prey survival was >99% under all control treatments and thus any mortality was due to predation by P. clarkii, which was also frequently observed. Procambarus clarkii were observed in pilot experiments to feed upon chrinomid larvae by searching with their antennae and maxillipeds, contrastingly they fed upon D. magna in a filter feeding fashion by generating a flow via tail flicking behaviours.
All model terms significantly affected prey consumption by P. clarkii (Table
Model terms for all factors from GLM with a quasi-Poisson error distribution used to determine differences in prey consumption with regards to factors “prey species”, “density”, and “substrate”, using a Type 3 Anova and χ2 to report the effect size of a factor on the dependent variable.
| Model term | Chisq | df | p-value |
|---|---|---|---|
| Prey species | 14 | 1 | <0.001 |
| Density | 207713 | 8 | <0.001 |
| Substrate | 78 | 2 | <0.001 |
| Prey species * Density | 607 | 8 | <0.001 |
| Prey species* Substrate | 112 | 2 | <0.001 |
| Density * Substrate | 1556 | 16 | <0.001 |
| Prey species * Density * Substrate | 1681 | 16 | <0.001 |
All prey species and substrate treatments resulted in a significant Type II FR by P. clarkii (Table
The FRR shows that when predating on chironomid larvae the impact of P. clarkii is highest on sand substrate compared to gravel or no substrate (Fig.

Functional responses of Procambarus clarkii preying on chironomid larvae and Daphnia magna under different substrate treatments; no substrate (solid line), sand (dashed line), gravel (dotted line). Points indicate raw data distributions; no substrate (green), sand (blue), gravel (brown). Shaded areas are bootstrapped (n = 2000) 95% confidence intervals.

Functional response ratio (FRR) (a/h) of Procambarus clarkii preying on chironomid larvae and Daphnia magna under different substrate treatments; no substrate (green), sand (blue), gravel (brown).
First order terms and significance levels from logistic regression of the proportion of prey consumed against initial prey density, with FR Type, functional parameters (a, h, and 1/h), associated significance levels from Rogers’ random predator equation, bias accelerated and corrected 95% confidence intervals for a and h, and the functional response ratio (a/h) with regards to Procambarus clarkii predating upon chironomid larvae and Daphnia magna under different substrate types. a = attack rate; h = handling time; 1/h = maximum feeding estimate.
| Prey | Substrate | First order term (p-value) | FR type | a (p-value) | a 95% BCa CI | h (p-value) | h 95% BCa CI | Maximum Feeding Estimate (1/h) | Functional Response Ratio (a/h) |
|---|---|---|---|---|---|---|---|---|---|
| Chironomid larvae | None | -0.016, <0.001 | II | 9.51, <0.001 | 6.79 – 3.53 | 0.007, <0.001 | 0.007–0.008 | 136.9 | 3102.7 |
| Sand | -0.016, <0.001 | II | 14.54, <0.001 | 9.30 – 28.8 | 0.007, <0.001 | 0.007–0.008 | 129.8 | 1888.3 | |
| Gravel | -0.016, <0.001 | II | 12.40, <0001 | 8.79 – 19.08 | 0.007, <0.001 | 0.007–0.008 | 131.5 | 1631.5 | |
| D. magna | None | -0.022, <0.001 | II | 8.37, <0.001 | 6.43 – 12.09 | 0.003, <0.001 | 0.003–0.004 | 263.1 | 2202.6 |
| Sand | -0.017, <0.001 | II | 14.50, <0.001 | 8.60 – 28.58 | 0.006, <0.001 | 0.006–0.007 | 144.9 | 2101.4 | |
| Gravel | -0.014, <0.001 | II | 9.63, <0.001 | 6.58 – 15.31 | 0.009, <0.001 | 0.008–0.009 | 111.1 | 1070.9 |
Discussion
Management of invasive species depends upon generating contextually relevant and accurate estimates of potential impact conveyed upon a recipient ecosystem (
Through the comparison of FR curves, it was possible to parameterise attack rate and handling time of a consumer upon a resource. Further, by comparing the FR curves for each prey species, we were able to distinguish whether one species may have higher pressures exerted upon it by an invasive species. Our results corroborate that P. clarkii indeed utilises both benthic and pelagic resources (
The presence of different substrata affects the predation efficiency of crayfish towards each prey type. Compared to the control treatment of no substrate, both sand, and gravel substrate caused a slight reduction in the attack parameter (or search efficiency) of P. clarkii upon the benthic chironomid prey rather than altering the handling time. This suggests that the presence of both sand and gravel can offer a refuge for benthic prey species as the crayfish is not able to access the meiofauna as readily. Similar trends are seen in benthic invasive gobiids, where gravel substrate reduces magnitude FR compared to sand substrates (
Daphnia magna is an important prey species for larval and adult crayfish (
Crayfish were able to generate a flow wherein the prey items were drawn closer, perhaps facilitating the low handling time demonstrated and indicating that filter feeding efficiency by adults is comparable to benthic foraging and thus warrants further investigation. Plasticity in feeding mode and flexible omnivory have been identified in other crustacean invaders (
Procambarus clarkii is a global polytrophic keystone consumer which has facilitated its spread and pervasion into numerous systems (
In the context of furthering approaches to impact prediction in invasion science, we establish here how the application of FR and FRR methods can be used to powerfully predict impact where traditional assessment methods would not have identified a difference between contexts. Furthermore, we discern that there is high potential for crayfish to differentially consume both benthic and pelagic macroinvertebrate species which has implications for nutrient cycling and resource provisioning for native species. In a destructive and spreading species like P. clarkii it is imperative to continue to determine scenarios wherein its predatory effect can be dampened in order to implement mitigation strategies as removal of crayfish once established is unfeasible.
Acknowledgements
We acknowledge the infrastructure and equipment provided by the NRF-SAIAB Research Platform and the funding channelled through the NRF-SAIAB Institutional Support system as well as the support received through the National Research Foundation – South African Research Chairs Initiative of the Department of Science and Technology (Inland Fisheries and Freshwater Ecology, Grant No. 110507). We thank the Queen’s University Marine Laboratory for use of laboratory facilities and a running grant.
References
- Alexander ME, Dick JTA, Weyl OLF, Robinson TB, Richardson DM (2014) Existing and emerging high impact invasive species are characterized by higher functional responses than natives. Biology Letters 10: e20130946. https://doi.org/10.1098/rsbl.2013.0946
- Aquiloni L, Ilhéu M, Gherardi F (2005) Habitat use and dispersal of the invasive crayfish Procambarus clarkii in ephemeral water bodies of Portugal. Marine and Freshwater Behaviour and Physiology 38: 225–236. https://doi.org/10.1080/10236240500310195
- Barrios-O’Neill D, Dick JTA, Emmerson MC, Ricciardi A, MacIsaac HJ (2015) Predator-free space, functional responses and biological invasions. Functional Ecology 29: 377–384. https://doi.org/10.1111/1365-2435.12347
- Bellard C, Cassey P, Blackburn TM (2016) Alien species as a driver of recent extinctions. Biology Letters 12: 20150623. https://doi.org/10.1098/rsbl.2015.0623
- Blackburn TM, Essl F, Evans T, Hulme PE, Jeschke JM, Kühn I, Kumschick S, Marková Z, Mrugała A, Nentwig W, Pergl J, Pyšek P, Rabitsch W, Ricciardi A, Richardson DM, Sendek A, Vilà M, Wilson JRU, Winter M, Genovesi P, Bacher S (2014) A unified classification of alien species based on the magnitude of their environmental impacts. PLOS Biology 12: e1001850. https://doi.org/10.1371/journal.pbio.1001850
- Bolker BM (2008) emdbook: ecological models and data in R. Princeton University Press, Princeton.
- Bolker BM (2010) bbmle: Tools for General Maximum Likelihood Estimation. R Package. http://www.cran.rproject.org
- Brown KM (1998) The role of shell strength in selective foraging by crayfish for gastropod prey. Freshwater Biology 40: 255–260. https://doi.org/10.1046/j.1365-2427.1998.00350.x
- Brown PB, Wetzel JE, Spacie A, Konopka A (1992) Evaluation of naturally-occurring organisms as food for juvenile crayfish Procambarus clarkii. Journal of the World Aquaculture Society 23: 211–216. https://doi.org/10.1111/j.1749-7345.1992.tb00771.x
- Bucciarelli GM, Suh D, Lamb AD, Roberts D, Sharpton D, Shaffer HB, Fisher RN, Kats LB (2019) Assessing effects of non-native crayfish on mosquito survival. Conservation Biology 33: 122–131. https://doi.org/10.1111/cobi.13198
- Budd TW, Lewis JC, Tracey ML (1978) The filter-feeding apparatus in crayfish. Canadian Journal of Zoology 56: 695–707. https://doi.org/10.1139/z78-097
- Carlton J, Fowler A (2018) Ocean rafting and marine debris: a broader vector menu requires a greater appetite for invasion biology research support. Aquatic Invasions 13: 11–15. https://doi.org/10.3391/ai.2018.13.1.02
- Carreira BM, Segurado P, Laurila A, Rebelo R (2017) Can heat waves change the trophic role of the world’s most invasive crayfish? Diet shifts in Procambarus clarkii. PLOS ONE 12: e0183108. https://doi.org/10.1371/journal.pone.0183108
- Chucholl C (2013) Invaders for sale: trade and determinants of introduction of ornamental freshwater crayfish. Biological Invasions 15: 125–141. https://doi.org/10.1007/s10530-012-0273-2
- Correia AM (2002) Niche breadth and trophic diversity: feeding behaviour of the red swamp crayfish (Procambarus clarkii) towards environmental availability of aquatic macroinvertebrates in a rice field (Portugal). Acta Oecologica 23: 421–429. https://doi.org/10.1016/S1146-609X(02)01166-9
- Courchamp F, Fournier A, Bellard C, Bertelsmeier C, Bonnaud E, Jeschke JM, Russell JC (2017) Invasion biology: specific problems and possible solutions. Trends in Ecology and Evolution 32: 13–22. https://doi.org/10.1016/j.tree.2016.11.001
- Cuthbert RN, Dalu T, Wasserman RJ, Dick JTA, Mofu L, Callaghan A, Weyl OLF (2018) Intermediate predator naïveté and sex-skewed vulnerability predict the impact of an invasive higher predator. Scientific Reports 8: 14282. https://doi.org/10.1038/s41598-018-32728-0
- Cuthbert RN, Dickey JWE, Coughlan NE, Joyce PWS, Dick JTA (2019) The Functional Response Ratio (FRR): advancing comparative metrics for predicting the ecological impacts of invasive alien species. Biological Invasions 21: 2543–2547. https://doi.org/10.1007/s10530-019-02002-z
- Davis AJS, Darling JA (2017) Recreational freshwater fishing drives non-native aquatic species richness patterns at a continental scale. Diversity and Distributions 23: 692–702. https://doi.org/10.1111/ddi.12557
- Dick JTA, Alexander ME, Jeschke JM, Ricciardi A, MacIsaac HJ, Robinson TB, Kumschick S, Weyl OLF, Dunn AM, Hatcher MJ, Paterson RA, Farnsworth KD, Richardson DM (2014) Advancing impact prediction and hypothesis testing in invasion ecology using a comparative functional response approach. Biological Invasions 16: 735–753. https://doi.org/10.1007/s10530-013-0550-8
- Dick JTA, Alexander ME, Ricciardi A, Laverty C, Downey PO, Xu M, Jeschke JM, Saul W-C, Hill MP, Wasserman R, Barrios-O’Neill D, Weyl OLF, Shaw RH (2017a) Functional responses can unify invasion ecology. Biological Invasions 19: 1667–1672. https://doi.org/10.1007/s10530-016-1355-3
- Dick JTA, Laverty C, Lennon JJ, Barrios-O’Neill D, Mensink PJ, Britton JR, Médoc V, Boets P, Alexander ME, Taylor NG, Dunn AM, Hatcher MJ, Rosewarne PJ, Crookes S, MacIsaac HJ, Xu M, Ricciardi A, Wasserman RJ, Ellender BR, Weyl OLF, Lucy FE, Banks PB, Dodd JA, MacNeil C, Penk MR, Aldridge DC, Caffrey JM (2017b) Invader Relative Impact Potential: a new metric to understand and predict the ecological impacts of existing, emerging and future invasive alien species. Journal of Applied Ecology 54: 1259–1267. https://doi.org/10.1111/1365-2664.12849
- Dickey JWE, Cuthbert RN, Rea M, Laverty C, Crane K, South J, Briski E, Chang X, Coughlan NE, MacIsaac HJ, Ricciardi A, Riddell GE, Xu M, Dick JTA (2018) Assessing the relative potential ecological impacts and invasion risks of emerging and future invasive alien species. NeoBiota 40: 1–24. https://doi.org/10.3897/neobiota.40.28519
- Evangelista C, Olden JD, Lecerf A, Cucherousset J (2019) Scale-dependent patterns of intraspecific trait variations in two globally invasive species. Oecologia 189: 1083–1094. https://doi.org/10.1007/s00442-019-04374-4
- Gebauer R, Veselý L, Vanina T, Buřič M, Kouba A, Drozd B (2019) Prediction of ecological impact of two alien gobiids in habitat structures of differing complexity. Canadian Journal of Fisheries and Aquatic Sciences 76(11): 1954–1961. https://doi.org/10.1139/cjfas-2018-0346
- Geiger W, Otero M, Alcorlo P (2004) Feeding preferences and food selection of the red swamp crayfish, Procambarus clarkii, in habitats differing in food item diversity. Crustaceana 77: 435–453. https://doi.org/10.1163/1568540041643283
- Gerlach SA, Ekstrøm DK, Eckardt PB (1976) Filter feeding in the hermit crab. Oecologia 24: 257–264. https://doi.org/10.1007/BF00345477
- Hänfling B, Edwards F, Gherardi F (2011) Invasive alien Crustacea: dispersal, establishment, impact and control. BioControl 56: 573–595. https://doi.org/10.1007/s10526-011-9380-8
- Hobbs HH, Barr TC (1960) The origins and affinities of the troglobitic crayfishes of North America (Decapoda, Astacidae). I. The genus Cambarus. The American Midland Naturalist 64: 12–33. https://doi.org/10.2307/2422890
- Holling CS (1959) Some characteristics of simple types of predation and parasitism. The Canadian Entomologist 91: 385–398. https://doi.org/10.4039/Ent91385-7
- Horton PA, Rowan M, Webster KE, Peters RH (1979) Browsing and grazing by cladoceran filter feeders. Canadian Journal of Zoology 57: 206–212. https://doi.org/10.1139/z79-019
- Jackson MC, Evangelista C, Zhao T, Lecerf A, Britton JR, Cucherousset J (2017) Between-lake variation in the trophic ecology of an invasive crayfish. Freshwater Biology 62: 1501–1510. https://doi.org/10.1111/fwb.12957
- Jeschke JM, Kopp M, Tollrian R (2004) Consumer-food systems: why type I functional responses are exclusive to filter feeders. Biological Reviews Cambridge Philosophical Society 79: 337–349. https://doi.org/10.1017/S1464793103006286
- Johnston K, Robson BJ, Fairweather PG (2011) Trophic positions of omnivores are not always flexible: evidence from four species of freshwater crayfish. Austral Ecology 36: 269–279. https://doi.org/10.1111/j.1442-9993.2010.02147.x
- Juliano SA (2001) Nonlinear curve fitting: predation and functional response curves. In: Scheiner SM, Gurevitch J (Eds) Design and Analysis of Ecological Experiments. Oxford University Press, Oxford, 178–196.
- Kemp JS, Aldridge DC (2018) Comparative functional responses to explain the impact of sympatric invasive bivalves (Dreissena spp.) under different thermal regimes. Journal of Molluscan Studies 84: 175–181. https://doi.org/10.1093/mollus/eyy006
- Kestrup ÅM, Dick JTA, Ricciardi A (2011) Interactions between invasive and native crustaceans: differential functional responses of intraguild predators towards juvenile hetero-specifics. Biological Invasions 13: 731–737. https://doi.org/10.1007/s10530-010-9863-z
- Keyler TD, Hrabik TR, Mensinger AF, Rogers LS, Gorman OT (2019) Effect of light intensity and substrate type on siscowet lake trout (Salvelinus namaycush siscowet) predation on deepwater sculpin (Myoxocephalus thompsonii). Hydrobiologia 840: 77–88. https://doi.org/10.1007/s10750-019-3944-5
- Kim S-K, Guo Q, Jang I-K (2015) Effect of Biofloc on the survival and growth of the postlarvae of three penaeids (Litopenaeus vannamei, Fenneropenaeus chinensis, and Marsupenaeus japonicus) and their Biofloc feeding efficiencies, as related to the morphological structure of the third maxilliped. Journal of Crustacean Biology 35: 41–50. https://doi.org/10.1163/1937240X-00002304
- Klose K, Cooper SD (2012) Contrasting effects of an invasive crayfish (Procambarus clarkii) on two temperate stream communities. Freshwater Biology 57: 526–540. https://doi.org/10.1111/j.1365-2427.2011.02721.x
- Kouba A, Tíkal J, Císař P, Veselý L, Fořt M, Příborský J, Patoka J, Buřič M (2016) The significance of droughts for hyporheic dwellers: evidence from freshwater crayfish. Scientific Reports 6: 26569. https://doi.org/10.1038/srep26569
- Kozák P, Ďuriš Z, Petrusek A, Buřič M, Horká I, Kouba A, Kozubíková-Balcarová E, Policar T, Němečková K (2015) Crayfish Biology and Culture. University of South Bohemia in České Budějovice, Faculty of Fisheries and Protection of Waters, Vodňany, 201–249.
- Langer SM, Weiss LC, Ekvall MT, Bianco G, Hansson L-A, Tollrian R (2019) A three-dimensional perspective of Daphnia’s swimming behavior with and without predator cues. Limnology and Oceanography 64: 1515–1525. https://doi.org/10.1002/lno.11132
- Larson ER, Twardochleb LA, Olden JD (2017) Comparison of trophic function between the globally invasive crayfishes Pacifastacus leniusculus and Procambarus clarkii. Limnology 18: 275–286. https://doi.org/10.1007/s10201-016-0505-8
- Laverty C, Green KD, Dick JTA, Barrios-O’Neill D, Mensink PJ, Médoc V, Spataro T, Caffrey JM, Lucy FE, Boets P, Britton JR, Pegg J, Gallagher C (2017) Assessing the ecological impacts of invasive species based on their functional responses and abundances. Biological Invasions 19: 1653–1665. https://doi.org/10.1007/s10530-017-1378-4
- Lipták B, Veselý L, Ercoli F, Bláha M, Buřič M, Ruokonen T, Kouba A (2019) Trophic role of marbled crayfish in a lentic freshwater ecosystem. Aquatic Invasions: 14: 299–309. https://doi.org/10.3391/ai.2019.14.2.09
- Lodge DM, Rosenthal SK, Mavuti KM, Muohi W, Ochieng P, Stevens SS, Mungai BN, Mkoji GM (2005) Louisiana crayfish (Procambarus clarkii) (Crustacea: Cambaridae) in Kenyan ponds: non-target effects of a potential biological control agent for schistosomiasis. African Journal of Aquatic Science 30: 119–124. https://doi.org/10.2989/16085910509503845
- MacNeil C, Dick JTA, Elwood RW (1997) The trophic ecology of freshwater Gammarus spp. (Crustacea: Amphipoda): Problems and perspectives concerning the functional feeding group concept. Biological Reviews of the Cambridge Philosophical Society 72: 349–364. https://doi.org/10.1017/S0006323196005038
- Meakin CA, Qin JG, Mair GC (2009) Zooplankton predation and consumption by freshwater crayfish, Cherax tenuimanus, at different sizes and light conditions. Journal of Freshwater Ecology 24: 159–167. https://doi.org/10.1080/02705060.2009.9664276
- Meyerson LA, Carlton JT, Simberloff D, Lodge DM (2019) The growing peril of biological invasions. Frontiers in Ecology and the Environment 17: 191–191. https://doi.org/10.1002/fee.2036
- Momot WT (1995) Redefining the role of crayfish in aquatic ecosystems. Reviews in Fisheries Science 3: 33–63. https://doi.org/10.1080/10641269509388566
- Murdoch WW (1969) Switching in general predators: experiments on predator specificity and stability of prey populations. Ecological Monographs 39: 335–354. https://doi.org/10.2307/1942352
- Nishijima S, Nishikawa C, Miyashita T (2017) Habitat modification by invasive crayfish can facilitate its growth through enhanced food accessibility. BMC Ecology 17: 37. https://doi.org/10.1186/s12898-017-0147-7
- Nunes AL, Hoffman AC, Zengeya TA, Measey GJ, Weyl OLF (2017a) Red swamp crayfish, Procambarus clarkii, found in South Africa 22 years after attempted eradication. Aquatic Conservation: Marine and Freshwater Ecosystems 27: 1334–1340. https://doi.org/10.1002/aqc.2741
- Nunes AL, Zengeya TA, Measey GJ, Weyl OLF (2017b) Freshwater crayfish invasions in South Africa: past, present and potential future. African Journal of Aquatic Science 42: 309–323. https://doi.org/10.2989/16085914.2017.1405788
- Oficialdegui FJ, Clavero M, Sánchez MI, Green AJ, Boyero L, Michot TC, Klose K, Kawai T, Lejeusne C (2019) Unravelling the global invasion routes of a worldwide invader, the red swamp crayfish (Procambarus clarkii). Freshwater Biology 64: 1382–1400. https://doi.org/10.1111/fwb.13312
- Panksepp JB, Huber R (2004) Ethological analyses of crayfish behavior: a new invertebrate system for measuring the rewarding properties of psychostimulants. Behavioural Brain Research 153: 171–180. https://doi.org/10.1016/j.bbr.2003.11.014
- Patoka J, Kalous L, Kopecký O (2014) Risk assessment of the crayfish pet trade based on data from the Czech Republic. Biological Invasions 16: 2489–2494. https://doi.org/10.1007/s10530-014-0682-5
- Platvoet D, Van Der Velde G, Dick JTA, Li S (2009) Flexible omnivory in Dikerogammarus villosus (Sowinsky, 1894) (Amphipoda) – Amphipod Pilot Species Project (AMPIS) Report 5. Crustaceana 82: 703–720. https://doi.org/10.1163/156854009X423201
- Pritchard DW (2014) frair: a package for functional response analysis in R. GitHub. https://github.com/dpritchard/frair
- Pritchard DW, Paterson RA, Bovy HC, Barrios-O’Neill D (2017) Frair: an R package for fitting and comparing consumer functional responses. Methods in Ecology and Evolution 8: 1528–1534. https://doi.org/10.1111/2041-210X.12784
- R Core Team (2018) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. https://www.R-project.org/
- Ricciardi A, Cohen J (2007) The invasiveness of an introduced species does not predict its impact. Biological Invasions 9: 309–315. https://doi.org/10.1007/s10530-006-9034-4
- Ricciardi A, Hoopes MF, Marchetti MP, Lockwood JL (2013) Progress toward understanding the ecological impacts of nonnative species. Ecological Monographs 83: 263–282. https://doi.org/10.1890/13-0183.1
- Rogers D (1972) Random search and insect population models. Journal of Animal Ecology 41: 369–383. https://doi.org/10.2307/3474
- SCBD (2010) COP-10 Decision X / 2: Strategic Plan for Biological Diversity 2011–2020. Secretariat of the Convention on Biological Diversity, Nagoya.
- Schoener TW (1971) Theory of feeding strategies. Annual Review of Ecology and Systematics 2: 369–404. https://doi.org/10.1146/annurev.es.02.110171.002101
- Seebens H, Blackburn TM, Dyer EE, Genovesi P, Hulme PE, Jeschke JM, Pagad S, Pyšek P, Winter M, Arianoutsou M, Bacher S, Blasius B, Brundu G, Capinha C, Celesti-Grapow L, Dawson W, Dullinger S, Fuentes N, Jäger H, Kartesz J, Kenis M, Kreft H, Kühn I, Lenzner B, Liebhold A, Mosena A, Moser D, Nishino M, Pearman D, Pergl J, Rabitsch W, Rojas-Sandoval J, Roques A, Rorke S, Rossinelli S, Roy HE, Scalera R, Schindler S, Štajerová K, Tokarska-Guzik B, van Kleunen M, Walker K, Weigelt P, Yamanaka T, Essl F (2017) No saturation in the accumulation of alien species worldwide. Nature Communications 8: 14435. https://doi.org/10.1038/ncomms14435
- Solomon ME (1949) The natural control of animal populations. Journal of Animal Ecology 18: 1–35. https://doi.org/10.2307/1578
- South J, Dick JTA, McCard M, Barrios-O’Neill D, Anton A (2017) Predicting predatory impact of juvenile invasive lionfish (Pterois volitans) on a crustacean prey using functional response analysis: effects of temperature, habitat complexity and light regimes. Environmental Biology of Fishes 100: 1155–1165. https://doi.org/10.1007/s10641-017-0633-y
- South J, Welsh D, Anton A, Sigwart JD, Dick JTA (2018) Increasing temperature decreases the predatory effect of the intertidal shanny Lipophrys pholis on an amphipod prey. Journal of Fish Biology 92: 150–164. https://doi.org/10.1111/jfb.13500
- Souty-Grosset C, Anastácio PM, Aquiloni L, Banha F, Choquer J, Chucholl C, Tricarico E (2016) The red swamp crayfish Procambarus clarkii in Europe: impacts on aquatic ecosystems and human well-being. Limnologica 58: 78–93. https://doi.org/10.1016/j.limno.2016.03.003
- Tokeshi M, Arakaki S (2012) Habitat complexity in aquatic systems: fractals and beyond. Hydrobiologia 685: 27–47. https://doi.org/10.1007/s10750-011-0832-z
- Wasserman RJ, Alexander ME, Dalu T, Ellender BR, Kaiser H, Weyl OLF (2016) Using functional responses to quantify interaction effects among predators. Functional Ecology 30: 1988–1998. https://doi.org/10.1111/1365-2435.12682