Research Article |
|
Corresponding author: Suzanne T. E. Lommen ( suzannelommen@hotmail.com ) Academic editor: Franz Essl
© 2018 Suzanne T. E. Lommen, Eelke Jongejans, Melinda Leitsch-Vitalos, Barbara Tokarska-Guzik, Mihály Zalai, Heinz Müller-Schärer, Gerhard Karrer.
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Lommen STE, Jongejans E, Leitsch-Vitalos M, Tokarska-Guzik B, Zalai M, Müller-Schärer H, Karrer G (2018) Time to cut: population models reveal how to mow invasive common ragweed cost-effectively. NeoBiota 39: 53-78. https://doi.org/10.3897/neobiota.39.23398
|
Abstract
Roadsides are an important habitat for invasive common ragweed, Ambrosia artemisiifolia L., by facilitating seed dispersal. Reducing the size of roadside populations is therefore essential for confining this highly allergenic species. Here, we aim to determine the cost-effectiveness of mowing regimes varying in frequency and timing, by analysing population-level effects and underlying demographic processes. We constructed population models of A. artemisiifolia parameterised by demographic data for four unmanaged reference populations across Europe in two years. We integrated the effects of four experimental mowing regimes along Austrian road sides on plant performance traits of five years and experimental data on seed viability after cutting. All four experimental regimes reduced the projected intrinsic population growth rates (r) compared to the unmanaged controls by reducing plant height and seed viability, thereby counteracting increased size-dependent fecundity. The prevailing 2-cut regime in Austria (cutting during vegetative growth, here in June and just before seed ripening, here in September) performed least well and the reduction in r was mainly due to reduced seed viability after the second cut. The efficacy of the two best experimental regimes (alternative schemes for 2 or 3 cuts) was mainly due to cutting just before female flowering (here in August) by decreasing final adult plant height dramatically and thereby reducing seed numbers. Patterns were consistent across reference populations and years. Whether regimes reduced r below replacement level, however, varied per population, year and the survival rate of the seeds in the soil bank. Our model allowed projecting effects of five theoretical mowing regimes with untested combinations of cuts on r. By plotting r-cost relationships for all regimes, we identified the most cost-effective schemes for each cutting frequency (1–3 cuts). They all included the cut just before female flowering, highlighting the importance of cutting at this moment (here in August). Our work features i) the suitability of a modelling approach for the demography of an annual species with a seed bank, ii) the importance of seed viability in assessing mowing effects, iii) the use of population models in designing cost-effective mowing regimes.
Keywords
Annual plant, population model, cost-effective management, population growth rate, seed viability, soil seed bank
Introduction
Successful management of invasive plant populations requires a cost-effective reduction of their population size, sufficient to mitigate their negative impact (
Common ragweed (Ambrosia artemisiifolia L.) is a worldwide invasive plant which has rapidly expanded in Europe in the past decades (
A multi-aspect experiment testing alternative mowing regimes along Austrian roadsides in multiple years showed that adapting the timing and/or frequency is required to reduce the seed production of individual A. artemisiifolia plants (
Here, we address these gaps in order to understand the efficacy of the tested mowing regimes in limiting growth of A. artemisiifolia populations, link these to management costs and design additional cost-effective mowing regimes. Specifically, we ask how mowing frequency and timing affect population growth of A. artemisiifolia? To answer this question, we analysed i) how mowing frequency and timing affect vital rates of plants in roadside populations and how these effects vary from year to year, ii) how mowing affects seed viability and iii) how much each of these mowing effects contribute to changes in population growth in different populations across Europe in different years. We construct population models for the study system and parameterise these with newly-collected demographic data from geographically distant populations across Europe and by integrating several existing and new data on the effect of mowing. Finally, we simulate new theoretical mowing regimes and compare their cost-effectiveness to the experimentally tested ones.
Methods
Study species
Ambrosia artemisiifolia originates from central USA and has invaded a wide range of habitat types on most other continents (

Life cycle of the annual plant Ambrosia artemisiifolia. Months indicated are representative for populations in Europe where most new plants have established in June, have developed into seed-producing plants in September and then die while the seeds shed overwinter on or in the soil. The small loop represents (dormant) seeds that do not recruit but stay in the soil until the next year. The vital rates and corresponding parameters in the demographic model are listed for each period. The x represents new plant size in June, y the plant size in September, m the effect of mowing.
Data
Overview of data and approach
In order to parameterise all vital rates and integrate the effects of mowing (Fig.
Roadside mowing experiment
A 5-year mowing experiment was set up along roadsides at six locations in 2009 and in a seventh location in 2010 in Austria. All locations were already infested with A. artemisiifolia for an unknown period and managed by a regular mowing regime for road maintenance, comprising a cut around June (during vegetative growth of A. artemisiifolia) and in September (just before seed ripening of A. artemisiifolia). In summary, each location was divided into five blocks of 20 m length each and, at each location, each of five treatments was randomly assigned to one block. Apart from the untreated control, experimental treatments were yearly applied by road service maintenance teams and included the prevailing mowing regime in eastern Austria and three alternative mowing regimes varying in the timing and frequency of cutting (upper half of Table
Overview of the experimental (upper half of table) and theoretical (lower half) mowing treatments. Their treatment code (indicating the calendar months in which cuts were conducted), the number and timing of cuts (each × indicating a cut) are shown.
| Treatment code | # cuts | Timing of cuts | Remarks | Model equation used | ||||
| Last week June (vegetative growth) | Last week July (before peak male flowering) | 3rd week Aug (before peak female flowering) | 2nd week Sept (before seed ripening) | |||||
| Experimental | 0 | 0 | Uncut control treatment (treatment 1 in mowing experiment) | 1 | ||||
| 69 | 2 | × | × | Prevailing regime in eastern Austria (treatment 2 in mowing experiment) | 2 | |||
| 89 | 2 | × | × | Experimental new treatment (treatment 3 in mowing experiment) | 2 | |||
| 679 | 3 | × | × | × | Experimental new treatment (treatment 4 in mowing experiment) | 2 | ||
| 689 | 3 | × | × | × | Experimental new treatment (treatment 5 in mowing experiment) | 2 | ||
| Theoretical | 6 | 1 | × | Modelled as treatment 69 without September cut | 2 | |||
| 8 | 1 | × | Modelled as treatment 89 without September cut | 2 | ||||
| 67 | 2 | × | × | Modelled as treatment 679 without September cut | 2 | |||
| 68 | 2 | × | × | Modelled as treatment 689 without September cut | 2 | |||
| 9 | 1 | × | Modelled as control treatment 0 added with a September cut | 2 | ||||
Post-harvest seed quality experiment
The effects of cutting on seed viability were obtained from a post-harvest seed quality experiment (
Demographic survey
To serve as “reference” populations for the current mowing effect study, we selected A. artemisiifolia populations located in the same bioclimatic region as the Austrian sites of the mowing experiment (Continental or Pannonian), with the most similar habitat type possible (grasslands, since no unmanaged roadsides were available) and with plant heights covering the range of plant heights observed in the untreated controls of the mowing experiment. This yielded four populations located in Austria (AT), Hungary (HU), Italy (IT) and Poland (PL) (details in Suppl. material
Seed burial experiments
To obtain values of seed survival rates, we used three of the largest available data sets for burial experiments with yearly measurements on our study species to date (
Population models
Population models of unmanaged references
Our discrete-time population model describes a time step of a year, from October (after seeds have been shed and plants have died) to October. As, in our model, individuals only exist as seeds in October, our model could be seen as unstructured. However, most vital rates in the model describe individual performance of plants from June to October and are functions of the continuous size variables x (plant height in June) or y (plant height in September). Our model is hence similar to a periodic Integral Projection Model with size as a continuous state variable (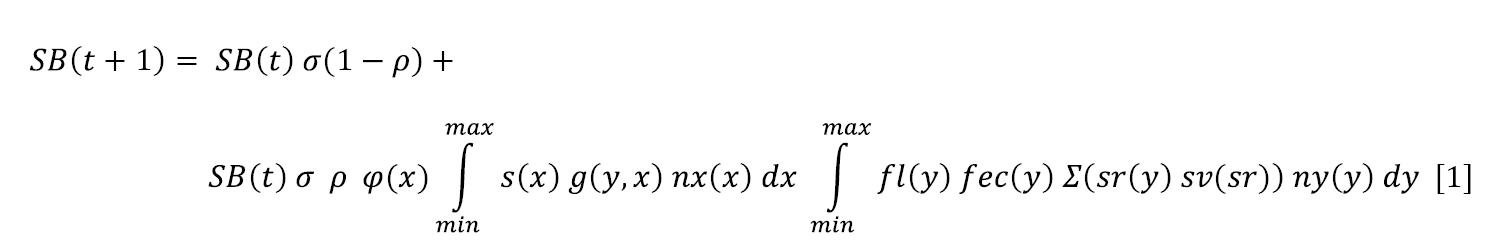
The first part of the equation describes the seeds that survived the entire year in the soil seed bank (small life cycle loop in Fig.
Using equation 1, we parameterised population models for all combinations of the six reference data sets and the three seed survival rates (i.e. a total of 18 reference scenarios, representing unmanaged controls). For details of the parameterisation, we refer to Suppl. material
Population models of experimental mowing treatments
To project the effects of the experimental mowing treatments, equation 1 was extended by including dependence on mowing treatment m into all the relevant vital rate functions, resulting in equation 2 (modifications to equation 1 are indicated in bold):
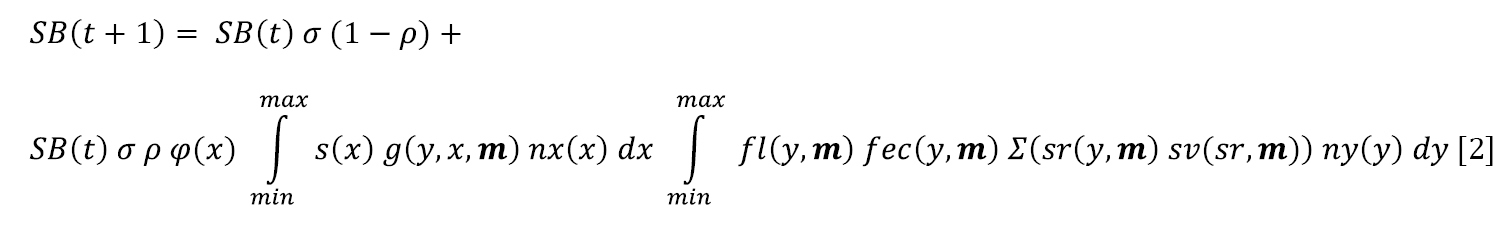
Details of the parameterisation are elaborated in Suppl. material
Simulating new theoretical mowing treatments with population models
Our approach allowed assessing the effect of new, experimentally untested combinations of cutting dates. Since the cut in September only affects population growth by modifying seed quality (through reduced seed ripening and corresponding lower seed viability) and no other vital rates, we were able to theoretically simulate removal or addition of this cut from the experimentally tested mowing regimes (Table
Analyses of population models
We obtained the projected intrinsic population growth rate, r, for each population model parameterised and compared them to assess the effect of treatments and the contribution of single vital rates. Since the effect of seed ripening only exerts an effect on r through the corresponding reduction in seed viability, we calculated the contribution of seed ripening alone as the difference between the growth rate when seed viability only was integrated and the growth rate when integrating both seed ripening and viability. To acknowledge temporal variation in experimental mowing effects (
Results
Effects of mowing on vital rates of plants
For plant height in September, the full model with the interaction of treatment and year fitted the data best. The pattern of treatment effects was nevertheless fairly consistent across the years without a clear pattern over time (Fig.

Fitted effects of experimental mowing treatments on vital rates. Experimental mowing treatments are indicated by their code (69, 89, 679, 689, see Table
Effects of mowing on seed quality
The best model for seed ripening contained the interaction between treatment and plant height. We found that mowing treatments generally delayed the development of seeds, especially for the transition from flowers to unripe seeds in smaller plants (Fig.
Effects of mowing on population growth
We projected stochastic intrinsic population growth, rs, for all 360 integrations of experimental mowing treatments and reference models. Although different seed survival scenarios resulted in very different values of r (Suppl. material

Effect of experimental mowing treatments on stochastic population growth rates rs for seed scenario H. Panels represent different populations (AT, HU, IT, PL) in year 2014 or 2015 (14 and 15, respectively) as detailed in Suppl. material
Contribution of vital rates and seed quality to population growth
We assessed the contribution of all vital rates and seed quality to changes in projected deterministic intrinsic population growth r, which were independent of seed survival. All treatments reduced r mainly through the decrease in growth (i.e. lower plant height in September) and seed viability counteracting the increased size-dependent seed production (Fig.

The influence of single vital rates and the September cut on population growth. Figures show the relative contributions of each vital rate (g = growth, fl = flowering, fec = fecundity, sr = seed ripening, sv = seed viability, see Fig.
Cost-effectiveness of mowing regimes
We plotted r-cost relationships of all experimental and theoretical mowing treatments, assuming that each cutting intervention has the same costs. Therefore, the relative costs are represented by the number of cuts. Fig.

Effect-cost relationships of mowing regimes for the Austrian reference population in 2014 (AT14) for seed survival scenario H. For each mowing regime (dots with labels indicating the months of cutting, see Table
Discussion
We show that optimising both the frequency and timing of cuts is the key to achieving the largest reductions in population growth rates (r) of A. artemisiifolia of roadsides by mowing. An increased frequency of cuts does not necessarily improve the effect, as was also found when mowing invasive thistles in the field (
We have provided a mechanistic understanding of how the experimental mowing regimes change projected population growth rates (Fig.
Our population modelling approach also unveiled the value of cuts at specific times through their effect on vital rates. Cutting during vegetative growth (here in June) has a small effect on final plant height (here in September) and the direction of the effect varied with year. Indeed, the species is known for its large regrowth capacity (
The results have wider implications for designing management strategies by using population models. A common approach is the identification of key life-cycle stages having the greatest impact on population growth rates (
Our population models are not meant to predict absolute values of intrinsic population growth. Firstly, we have shown that the reported r values highly depend on the persistence in the soil seed bank. Our best estimates for seed survival came from burial experiments, while in roadside populations, seeds are unlikely to be buried deeply. When they remain on the surface of the ground, they are exposed to different abiotic conditions and other factors that may cause additional mortality, such as seed predation. Our scenarios for seed survival are therefore conservative. Although the pattern of the r-cost curves is independent of seed survival, mowing will bring r to lower values when seed survival is lower. It is known in other systems, especially annuals with a seed bank, that demographic models can be strongly influenced by seed survival rates (
Our r-cost chart shows the relative costs, corresponding to the number of yearly cuts which a mowing regime comprises. The absolute costs for mowing along roadsides per cut per kilometre are unavailable. They depend on very specific circumstances of the responsible authorities. For instance, the number of workers and machinery needed at distinct dates for spatiotemporally fitted optimal mowing varies between countries and regions (personal observation, G. Karrer). In many cases, additional personnel and machinery have to be rented, adding costs to the fixed expenses for regular personnel and machinery.
We are aware that we used our models mainly to assess mowing effects on population size, while other results of local demography and treatments may be of interest to managers as well, such as the total seed output (discussed above), pollen production or population spread. Optimal management for reducing the number of plants does not necessarily need to coincide with optimal management of other target variables (
Our study focused on cost-effective local management of roadsides and evaluated management impact by population size of the target. For a comprehensive economic assessment of the efficacy of management of A. artemisiifolia at the regional level, however, efficacy beyond the population level should be assessed. As Ambrosia artemisiifolia occurs in different climatic areas (
Conclusions
Overall, our population modelling approach has proven to be a useful tool for comparing population-level effects of different mowing regimes for an annual plant with a long-lasting seed bank. Integration of mowing effects into reference models of four geographically distant populations in Europe in two different years showed that patterns of projected population effects were consistent across time and space (Suppl. material
Acknowledgements
We are thankful to all land owners for allowing us to undertake the demographic studies and to Maira Bonini and her team for arranging the Italian site. We greatly appreciate all the help in demographic data collection by Eva van Cleef, Simon Vandenbrande, Elisa Cardarelli, Silvia Stefanelli, Stéphanie von Bergen, Gaelle Kadima, Sanela Milenkovic, Aleksandra Krsmanovic, Nina Waldhäuser and Ivana Milakovic. We also thank an anonymous reviewer for constructive comments on cost-effectiveness. This research was funded by the Swiss State Secretariat for Education, Research and Innovation (C13.0146; to HMS), the Swiss Federal Office for the Environment (13.0098.KP/M323-0760; to HMS), the Netherlands Organisation for Scientific Research (NWO-grants 840.11.001/841.11.007; to EJ), the Austrian Ministry of Agriculture, Forestry, Environment and Water and eight Federal State Governments of Austria, the EFOP-3.6.3-VEKOP-16-2017-00008 project co-financed by the European Union and the European Social Fund, the Department of Botany and Nature Protection of the Faculty of Biology and Environmental Protection of the University of Silesia in Katowice (to BTG), DG Environment of the European Commission (project ‘HALT Ambrosia’, Grant Agreement No. 07.0322/2010/586340/SUB/B2) and the EU COST Action FA1203 ‘Sustainable management of Ambrosia artemisiifolia in Europe (SMARTER)’ (
References
- Barbour BM, Meade JA (1981) The effects of cutting date and height on anthesis of common ragweed (Ambrosia artemisiifolia L.). Proceedings of the Annual Meeting Northeast Weed Science Society 35: 82–86.
- Basky Z, Ladányi M, Simončič A (2017) Efficient reduction of biomass, seed and season long pollen production of common ragweed (Ambrosia artemisiifolia L.). Urban Forestry & Urban Greening 24: 134–140. https://doi.org/10.1016/j.ufug.2017.03.028
- Bassett IJ, Crompton CW (1975) The biology of Canadian weeds: Ambrosia artemisiifolia L. and A. psilostachya DC. Canadian Journal of Plant Science 55: 463–476. https://doi.org/10.4141/cjps75-072
- Bazzaz FA (1979) Physiological ecology of plant succession. Annual Review of Ecology and Systematics 10: 351–371. https://doi.org/10.1146/annurev.es.10.110179.002031
- Bohren C, Delabays N, Mermillod G, Baker A, Vertenten J (2008) Ambrosia artemisiifolia L: Optimieren des Schnittregimes. Agrarforschung 15: 308–313.
- Bourdôt GW, Basse B, Cripps MG (2016) Mowing strategies for controlling Cirsium arvense in a permanent pasture in New Zealand compared using a matrix model. Ecology and Evolution 6: 2968–2977. https://doi.org/10.1002/ece3.2090
- Buckley YM, Briese DT, Rees M (2003) Demography and management of the invasive plant species Hypericum perforatum. I. Using multi-level mixed-effects models for characterizing growth, survival and fecundity in a long-term data set. Journal of Applied Ecology 40: 481–493. https://doi.org/10.1046/j.1365-2664.2003.00821.x
- Buttenschøn RM, Waldispühl S, Bohren C (2009) Guidelines for management of common ragweed, Ambrosia artemisiifolia. University of Copenhagen, 53 pp.
- Caplat P, Coutts S, Buckley YM (2012) Modeling population dynamics, landscape structure, and management decisions for controlling the spread of invasive plants. In: Ostfeld RS, Schlesinger WH (Eds) Year in Ecology and Conservation Biology.Blackwell Science Publ, Oxford, 72–83. https://doi.org/10.1111/j.1749-6632.2011.06313.x
- Caswell H (1978) General formula for sensitivity of population growth rate to changes in life-history parameters Theoretical Population Biology 14: 215–230. https://doi.org/10.1016/0040-5809(78)90025-4
- Chapman DS, Makra L, Albertini R, Bonini M, Paldy A, Rodinkova V, Sikoparija B, Weryszko-Chmielewska E, Bullock JM (2016) Modelling the introduction and spread of non-native species: international trade and climate change drive ragweed invasion. Global Change Biology 22: 3067–3079. https://doi.org/10.1111/gcb.13220
- Crone EE, Ellis MM, Morris WF, Stanley A, Bell T, Bierzychudek P, EhrlÉN J, Kaye TN, Knight TM, Lesica P, Oostermeijer G, Quintana-Ascencio PF, Ticktin T, Valverde T, Williams JL, Doak DF, Ganesan R, McEachern K, Thorpe AS, Menges ES (2013) Ability of Matrix Models to Explain the Past and Predict the Future of Plant Populations. Conservation Biology 27: 968–978. https://doi.org/10.1111/cobi.12049
- Crone EE, Menges ES, Ellis MM, Bell T, Bierzychudek P, Ehrlén J, Kaye TN, Knight TM, Lesica P, Morris WF, Oostermeijer G, Quintana-Ascencio PF, Stanley A, Ticktin T, Valverde T, Williams JL (2011) How do plant ecologists use matrix population models? Ecology Letters 14: 1–8. https://doi.org/10.1111/j.1461-0248.2010.01540.x
- Domonkos Z, Szigeti Szabó V, Farkas A, Pinke G, Reisinger P, Vereš T, Tóth P (2017) Spread of common ragweed (Ambrosia artemisiifolia L.) on arable land in the Žitný ostrov. Journal of Central European Agriculture 18: 29–41. https://doi.org/10.5513/JCEA01/18.1.1863
- Ellner SP, Rees M (2006) Integral projection models for species with complex demography. American Naturalist 167: 410–428. https://doi.org/10.1086/499438
- Essl F, Biró K, Brandes D, Broennimann O, Bullock JM, Chapman DS, Chauvel B, Dullinger S, Fumanal B, Guisan A, Karrer G, Kazinczi G, Kueffer C, Laitung B, Lavoie C, Leitner M, Mang T, Moser D, Müller-Schärer H, Petitpierre B, Richter R, Schaffner U, Smith M, Starfinger U, Vautard R, Vogl G, von der Lippe M, Follak S (2015) Biological Flora of the British Isles: Ambrosia artemisiifolia. Journal of Ecology 103: 1069–1098. https://doi.org/10.1111/1365-2745.12424
- Essl F, Dullinger S, Kleinbauer I (2009) Changes in the spatio-temporal patterns and habitat preferences of Ambrosia artemisiifolia during its invasion of Austria. Preslia 81: 119–133.
- Gioria M, Pyšek P, Moravcova L (2012) Soil seed banks in plant invasions: promoting species invasiveness and long-term impact on plant community dynamics. Preslia 84: 327–350.
- Gross CL, Mackay D (2014) Two decades of demography reveals that seed and seedling transitions limit population persistence in a translocated shrub. Annals of Botany 114: 85–96. https://doi.org/10.1093/aob/mcu082
- Hansen MJ (2007) Evaluating management strategies and recovery of an invasive grass (Agropyron cristatum) using matrix population models. Biological Conservation 140: 91–99. https://doi.org/10.1016/j.biocon.2007.07.028
- Kalisz S, McPeek MA (1992) Demography of an age-structured annual: resampled projection matrices, elasticity analyses, and seed bank effects. Ecology 73: 1082–1093. https://doi.org/10.2307/1940182
- Kalisz S, McPeek MA (1993) Extinction dynamics, population growth and seed banks. Oecologia 95: 314–320. https://doi.org/10.1007/bf00320982
- Karrer G (2016a) Field experiment on longevity of the seeds in the soil seed bank (initial seed burial experiment at the University of Natural Resources and Life Sciences BOKU). In: Sölter U, Starfinger U, Verschwele A (Eds) HALT Ambrosia – final project report and general publication of project findings (Julius-Kühn-Archiv 455).Quedlinburg, 48–49. https://doi.org/10.5073/jka.2016.455.18
- Karrer G (2016b) Implications of life history for control and eradication. In: Söltner U, Starfinger U, Verschwele A (Eds) HALT Ambrosia – final project report and general publication of project findings (Julius-Kühn-Archiv 455).Quedlinburg, 58–64. https://doi.org/10.5073/jka.2016.455.18
- Karrer G (2016c) Post harvest seed ripening (pot experiment). In: Sölter U, Starfinger U, Verschwele A (Eds) HALT Ambrosia – final project report and general publication of project findings (Julius-Kühn-Archiv 455).Quedlinburg, 37–40. https://doi.org/10.5073/jka.2016.455.18
- Karrer G, Hall R, Lener F, Waldhäuser N, Kazinczi G, Kerepesi I, Máté S, Sölter U, Starfinger U, Verschwele A, Mathiassen SK, Kudsk P, Leskovšek R, Simončič A (2016) Field experiment on longevity of the seeds in the soil seed bank (Joint experiment). In: Sölter U, Starfinger U, Verschwele A (Eds) HALT Ambrosia – final project report and general publication of project findings (Julius-Kühn-Archiv 455).Quedlinburg, 41–47. https://doi.org/10.5073/jka.2016.455.18
- Karrer G, Milakovic M, Kropf M, Hackl G, Essl F, Hauser M, Mayer M, Blöch C, Leitsch-Vitalos M, Dlugosch A, Hackl G, Follak S, Fertsak S, Schwab M, Baumgarten A, Gansberger M, Moosbeckhofer R, Reiter E, Publig E, Moser D, Kleinbauer I, Dullinger S (2011) Dispersal and management of a highly allergenic introduced plant – pathways and causes for the dispersal of ragweed (Ambrosia artemisiifolia) and options for control. Vienna, 329 pp.
- Kazinczi G, Novák R, Pathy Z, Béres I (2008) Common ragweed (Ambrosia artemisiifolia L.): a review with special regards to the results in Hungary. III. Resistant biotypes, control methods and authority arrangements. Herbologia: an international journal on weed research and control 9: 119–144.
- Kerr NZ, Baxter PWJ, Salguero-Gómez R, Wardle GM, Buckley YM (2016) Prioritizing management actions for invasive populations using cost, efficacy, demography and expert opinion for 14 plant species world-wide. Journal of Applied Ecology 53: 305–316. https://doi.org/10.1111/1365-2664.12592
- Lennartsson T, Oostermeijer JGB (2001) Demographic variation and population viability in Gentianella campestris: effects of grassland management and environmental stochasticity. Journal of Ecology 89: 451–463. https://doi.org/10.1046/j.1365-2745.2001.00566.x
- Lommen STE, Hallmann CA, Jongejans E, Chauvel B, Leitsch-Vitalos M, Aleksanyan A, Tóth P, Preda C, Šćepanović M, Onen H, Tokarska-Guzik B, Anastasiu P, Dorner Z, Fenesi A, Karrer G, Nagy K, Pinke G, Tiborcz V, Zagyvai G, Zalai M, Kazinczi G, Leskovšek R, Stešević D, Fried G, Kalatozishvili L, Lemke A, Müller-Schärer H (2017) Explaining variability in the production of seed and allergenic pollen by invasive Ambrosia artemisiifolia across Europe. Biological Invasions. https://doi.org/10.1007/s10530-017-1640-9
- Metcalf CJE, Ellner SP, Childs DZ, Salguero-Gomez R, Merow C, McMahon SM, Jongejans E, Rees M (2015) Statistical modelling of annual variation for inference on stochastic population dynamics using Integral Projection Models. Methods in Ecology and Evolution 6: 1007–1017. https://doi.org/10.1111/2041-210x.12405
- Metcalf CJE, McMahon SM, Salguero-Gomez R, Jongejans E (2013) IPMpack: an R package for integral projection models. Methods in Ecology and Evolution 4: 195–200. https://doi.org/10.1111/2041-210x.12001
- Milakovic I, Fiedler K, Karrer G (2014a) Fine-tuning of a mowing regime, a method for the management of the invasive plant, Ambrosia artemisiifolia, at different population densities. Weed Biology and Management 14: 232–241. https://doi.org/10.1111/wbm.12051
- Milakovic I, Fiedler K, Karrer G (2014b) Management of roadside populations of invasive Ambrosia artemisiifolia by mowing. Weed Research 54: 256–264. https://doi.org/10.1111/wre.12074
- Milakovic I, Karrer G (2016) The influence of mowing regime on the soil seed bank of the invasive plant Ambrosia artemisiifolia L. NeoBiota 28: 39–49. https://doi.org/10.3897/neobiota.28.6838
- Müller-Schärer H, Lommen S (2014) EU-COST Action on „Sustainable management of Ambrosia artemisiifolia in Europe“ (COST FA1203-SMARTER): opportunities and challenges. Julius-Kühn-Archiv 445 148–155. https://doi.org/10.5073/jka.2013.445.018
- Normand S, Zimmermann NE, Schurr FM, Lischke H (2014) Demography as the basis for understanding and predicting range dynamics. Ecography 37: 1149–1154. doi:https://doi.org/10.1111/ecog.01490
- Patracchini C, Vidotto F, Ferrero A (2011) Common ragweed (Ambrosia artemisiifolia) growth as affected by plant density and clipping. Weed Technology 25: 268–276. https://doi.org/10.1614/wt-d-09-00070.1
- R Core Team (2017) R: A language and environment for statistical computing http://www.R-project.org/ [accessed 10 March.2017]
- Ramula S, Knight TM, Burns JH, Buckley YM (2008) General guidelines for invasive plant management based on comparative demography of invasive and native plant populations. Journal of Applied Ecology 45: 1124–1133. https://doi.org/10.1111/j.1365-2664.2008.01502.x
- Richter R, Berger UE, Dullinger S, Essl F, Leitner M, Smith M, Vogl G, Firn J (2013a) Spread of invasive ragweed: climate change, management and how to reduce allergy costs. Journal of Applied Ecology 50: 1422–1430. doi:https://doi.org/10.1111/1365-2664.12156
- Richter R, Dullinger S, Essl F, Leitner M, Vogl G (2013b) How to account for habitat suitability in weed management programmes? Biological Invasions 15: 657–669. https://doi.org/10.1007/s10530-012-0316-8
- Salguero-Gómez R, De Kroon H (2010) Matrix projection models meet variation in the real world. Journal of Ecology 98: 250–254. https://doi.org/10.1111/j.1365-2745.2009.01635.x
- Schindler S, Bayliss HR, Essl F, Rabitsch W, Follak S, Pullin AS (2016) Effectiveness of management interventions for control of invasive common ragweed Ambrosia artemisiifolia: a systematic review protocol. Environmental Evidence 5: 11. https://doi.org/10.1186/s13750-016-0062-y
- Shea K, Jongejans E, Skarpaas O, Kelly D, Sheppard AW (2010) Optimal management strategies to control local population growth or population spread may not be the same. Ecological Applications 20: 1148–1161. https://doi.org/10.1890/09-0316.1
- Shea K, Kelly D (1998) Estimating biocontrol agent impact with matrix models: Carduus nutans in New Zealand. Ecological Applications 8: 824–832. https://doi.org/10.2307/2641269
- Simard M-J, Benoit DL (2011) Effect of repetitive mowing on common ragweed (Ambrosia artemisiifolia L.) pollen and seed production. Annals of Agricultural and Environmental Medicine 18: 55–62.
- Simberloff D (2003) How much information on population biology is needed to manage introduced species? Conservation Biology 17: 83–92. https://doi.org/10.1046/j.1523-1739.2003.02028.x
- Skálová H, Guo W-Y, Wild J, Pyšek P (2017) Ambrosia artemisiifolia in the Czech Republic: history of invasion, current distribution and prediction of future spread. Preslia 89: 1–16. https://doi.org/10.23855/preslia.2017.001
- Smith M, Cecchi L, Skjoth CA, Karrer G, Sikoparija B (2013) Common ragweed: a threat to environmental health in Europe. Environment international 61: 115–126. https://doi.org/10.1016/j.envint.2013.08.005
- Sölter U, Verschwele A, Starfinger U (2016) Viability of seeds ripened after cutting (pot experiment). In: Sölter U, Starfinger U, Verschwele A (Eds) HALT Ambrosia – final project report and general publication of project findings (Julius-Kühn-Archiv 455). Quedlinburg, 36. https://doi.org/10.5073/jka.2016.455.12
- Sun Y, Broennimann O, Roderick G, Poltavsky A, Lommen STE, Mueller-Schaerer H (2017) Climatic suitability ranking of biological control candidates: a biogeographic approach for ragweed management in Europe. Ecosphere.
- Tokarska-Guzik B, Bzdęga K, Koszela K, Żabińska I, Krzuś B, Sajan M, Sendek A (2011) Allergenic invasive plant Ambrosia artemisiifolia L. in Poland: threat and selected aspects of biology. Biodiversity: Research and Conservation, 39 pp. https://doi.org/10.2478/v10119-011-0008-8
- Toole E, Brown E (1946) Final results of the Durvel buried seed experiment. Journal of Agricultural Research 72: 201–210.
- Vitalos M, Karrer G (2009) Dispersal of Ambrosia artemisiifolia seeds along roads: the contribution of traffic and mowing machines. In: Pyšek P, Pergl J (Eds) Biological Invasions: Towards a Synthesis, Proceedings of the 5th Neobiota Conference, September 2008.Inst. of Ecology of the TU Berlin, Prague, Czech Republic, 53–60.
- von der Lippe M, Bullock JM, Kowarik I, Knopp T, Wichmann MC (2013) Human-mediated dispersal of seeds by the airflow of vehicles. Plos ONE 8: e52733. https://doi.org/10.1371/journal.pone.0052733
- Williams JL, Miller TEX, Ellner SP (2012) Avoiding unintentional eviction from integral projection models. Ecology 93: 2008–2014. https://doi.org/10.1890/11-2147.1
- Zalai M, Poczok L, Dorner Z, Körösi K, Pálinkás Z, Szalai M, O. P (2017) Developing control strategies against common milkweed (Asclepias syriaca L.) on ruderal habitats. Herbologia 16: 69–84. https://doi.org/10.5644/Herb.16.2.07
- Zhang R, Shea K (2012) Integrating multiple disturbance aspects: management of an invasive thistle, Carduus nutans. Annals of Botany 110: 1395–1401. https://doi.org/10.1093/aob/mcr312
Supplementary materials
Analysis of mowing experiment data (statistical analysis of empirical data)
Demographic survey of reference populations (location table, methods)
Burial experiments (location table, methods, graphic results)
Parametrisation of population models of unmanaged references (model parameterisation)
Parametrisation of population models of experimental mowing treatments (model parameterisation)
Stochastic population growth for alternative seed bank scenarios (graphic results, population dynamics)
Deterministic population models per reference data set (graphic results, population dynamics)
r-cost curves for different seed survival scenarios and reference data sets (graphic results, population dynamics)